|
|
Митохондриальная наследственность
Дрожжи занимают центральное место в исследованиях по генетике митохондрий.
Жизненный цикл основного штамма дрожжей (Sаcсharоmyces cerevisiae), используемого в генетических исследованиях, весьма несложен. Дифференциа-ция гаплоидных штаммов определяется парой хромосомных аллелей а и 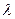 , контролирующих типы скрещиваемости. При смешивании в определенных условиях отдельные клетки а и сливаются между собой, образуя диплоидные зиготы, которые могут затем делиться путем митоза и давать начало клонам диплоидных клеток. У одиночных диплоидных клеток можно вызвать спору-ляцию, процесс, который связан с мейозом и образованием неупорядоченной тетрады, состоящей из аскоспор. В каждой тетраде аллели а и распределяют-ся в соотношении 2 : 2, и каждая спора затем при делении образует гаплоидный клон.
, контролирующих типы скрещиваемости. При смешивании в определенных условиях отдельные клетки а и сливаются между собой, образуя диплоидные зиготы, которые могут затем делиться путем митоза и давать начало клонам диплоидных клеток. У одиночных диплоидных клеток можно вызвать спору-ляцию, процесс, который связан с мейозом и образованием неупорядоченной тетрады, состоящей из аскоспор. В каждой тетраде аллели а и распределяют-ся в соотношении 2 : 2, и каждая спора затем при делении образует гаплоидный клон.
Особенности Sаcсharоmyces cerevisiae:
1. При росте на глюкозе обнаруживают уникальную способность суще-ствовать только за счет гликолиза и поэтому могут обходиться без функцио-нально активных митохондрий, т.е. без окислительного фосфорилирования. Это дает возможность разработать с клетками, митохондриальная и ядерная ДНК которых несут мутации, препятствующие нормальному развитию митохондрий. Такие мутации летальны почти у всех организмов.
2. Дрожжи – простые одноклеточные эукариоты, легко выращивать и подвергать биохимическим исследованиям.
3. Наряду с размножением путем почкования (асимметричного митоза) встречается и половой процесс.
Возможность контролировать в лабораторных условиях чередования бесполого и полового размножения намного облегчает проведение генетического анализа, который позволяет выявить гены, ответственные за функцию митохондрий и установить, которые из них находятся в ядерной ДНК, а какие в митохондриальной ДНК.
Образование в культуре пекарских дрожжей Sacchаromyces cerevisiaе (сахаромицетов) медленно растущих колоний, мелких колоний впервые было описано в 1949 г. Эфрусси. Эта мутация petite.
В результате биохимических исследований было установлено, что мутанты petite лишены способности к дыханию (аэробному окислению) из-за недостаточной активности дыхательных ферментов: цитохром оксидазы, цитохром-С-редуктазы, цитохромов а, в, а3, но содержат цитохром с. Этим объясняется медленный рост этих мутантов и рост только на сбраживаемых субстратах (например, глюкоза). Известно, что все эти ферменты локализуются в митохондриях, т.е. митохондрии клеток petite функционально неполноценны. Мутанты petite имеют аномалии в строении митохондрий.
Дрожжи, благодаря своей способности получать энергию с помощью другого метаболического процесса – брожения, могут расти в отсутствие кислорода. При выращивании в бескислородной среде разница в скорости роста нормальных штаммов и мутантов petite исчезает, они могут получать энергию с помощью брожения.
В отличие от нормальных штаммов мутанты petite не чувствительны к таким дыхательным ядам, как цианидам. Интенсивность брожения не зависит от наличия кислорода, оно не чувствительно к синильной кислоте.
Мутантные petite (реt-) возникают спонтанно гораздо чаще, чем большинство генных мутаций – » 1 мутация на 500 клеток за одну клеточную генерацию. Путем облучения УФ и обработки химическими мутагенами – красителями акридинового ряда, такими как эуфлавин, акрифлавин, бромистый этидий, частоту petite мутации можно повысить почти до 100%.
Генетический анализ мутантов petite дал следующие результаты:
Рe+ х pet- (дикий тип)
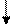
pet+ -диплоидные клетки, обладавшие подобно родителю дикого
 типа нормальным механизмом дыхания
типа нормальным механизмом дыхания
4 pet+ : 0 pet- -в результате споруляции этих диплоидных клеток
происходило образование аскоспор, которые все, за
редкими исключениями, оказывались нормальными.
Фенотип petite исчезал. Расщепления в тетрадах не обнаружилось. После этого из поколения F1 выбирали случайным образом клетки дикого типа и снова их скрещивали с первоначальным родителем реt-, все потомство было нормальным. Подобного рода возвратного скрещивания проводили на протяжении четырех поколений.
Вывод: результаты генетического анализа несовместимы с менделевским наследованием, мутация petite имеет цитоплазматическую природу (1949 г.)
За время поддержания первоначальных штаммов petite в культуре не было обнаружено ни одной реверсии. Данные штаммы называют нейтральными или рецессивными формами petite.
В 1955 г. был открыт другой класс petite – супрессивных или доминантных.
При скрещивании клеток некоторых мутантов pet- с клетками дикого типа с разной частотой (до 99 %) образуются диплоиды, не способные к дыханию. Это свойство, названное супрессивностью, стабильно наследовалось при вегетативном размножении и при скрещиваниях. Тетрадный анализ признака супресивности показывает отсутствие расщепления в подавляющем большинстве тетрад 4 рet- : 0 pet+.
Супрессивные и нейтральные мутанты – это вегетативные мутанты.
В 1950 г. Чен и др. сообщили о выделении ряда мутантов с фенотипом petite, но дающих менделевское расщепление при скрещивании:
Рet+х pet- (дикий тип)
 все диплоидные зиготы нормальные
все диплоидные зиготы нормальные
тетрада аскоспор, образующихся при споруляции, расщеплялась в соотношении 2 рet- : 2 pet+
Такие формы называли расщепляющимися (ядерными).
Найдено более 20 различных ядерных генов, большей частью несцеплен-ных, которые влияют на развитие митохондрий и обусловливают мутантный фенотип, сходный с фенотипом цитоплазматических форм petite. Это генеративныемутанты.
Скрещивания между ядерными и цитоплазматическими формами petite дают зиготу с нормальным дыханием, а после споруляции тетрада аскоспор расщепляется в соотношении 2 рet- : 2 нормальные.
В отличие от этого различные нейтральные формы petite