|
|
Вариабельность генома митохондрий
Структурно-функциональные свойства геномов митохондрий очень разнообразны в зависимости от таксономической принадлежности объекта.
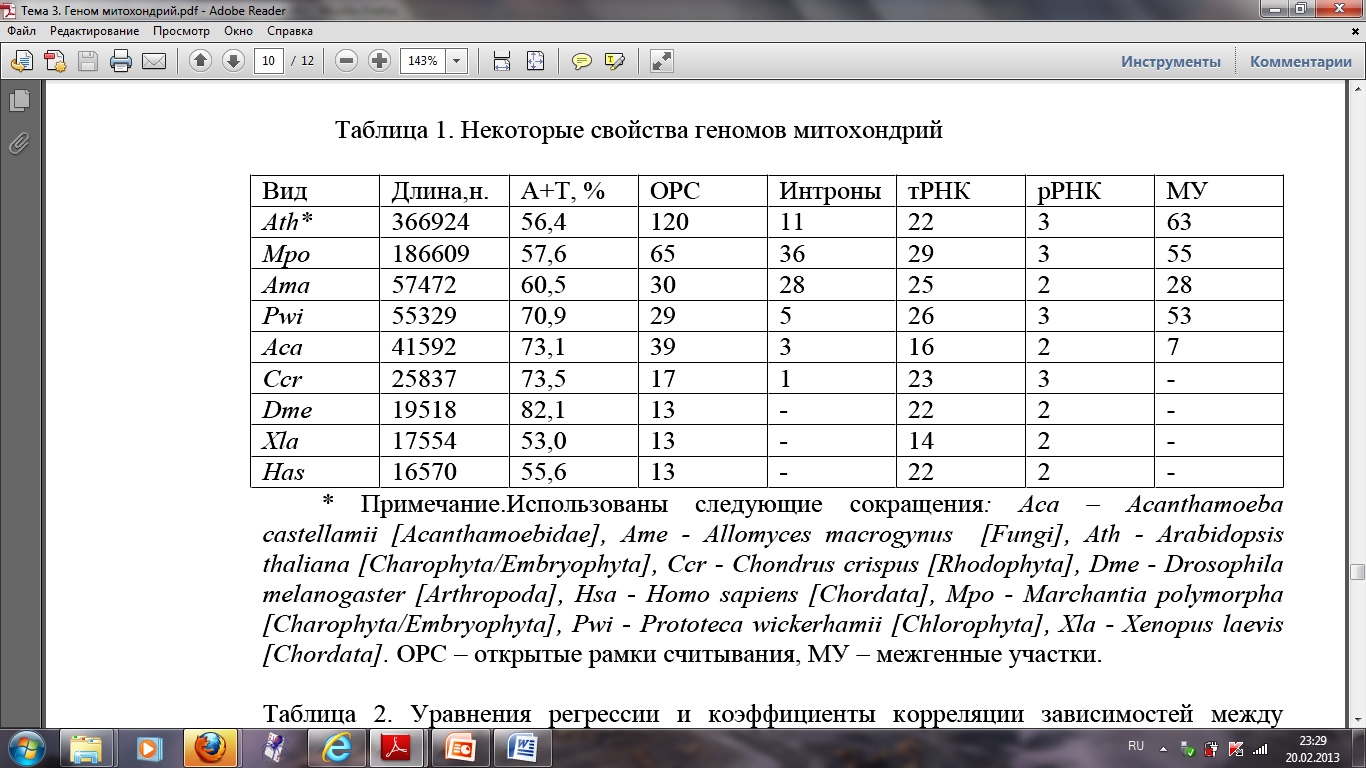

Число генов в ДНК митохондрий позвоночных и беспозвоночных практически постоянно и равно 36-37.
Типовой набор м-х-х генов включает набор генов:
1) кодирующих субъединицы комплекса I (NADH: убихинон оксидоредуктазы) nad 1–9;
2) комплекса III (убихинон:цитохром с оксидоредуктазы) cob;
3) комплекса IV (цитохром с оксидазы) cox 1–3;
4) комплекса V (ATP синтазы) atp 1,6,8.
5) гены рРНК большой и малой субчастиц рибосом
6) 22 гена тРНК;
7) 2 гена рРНК.
Этот набор генов за редким исключением присутствует практически во всех мт‑геномах и в частности в геномах первого варианта.
1)Геном митохондрий кл-к большинства жив-х– это кольцевая молекула размером 14–20 Kbp. Геном кодирует: 2 рРНК, 13 специфических полипептидов, до 25 тРНК (кол-во может меняться от 2 до 24 у разных видов), содержит незначительный процент некодирующей ДНК. Интроны в генах белков отсутствуют. Гены тРНК расположены м/у генами белков и, кроме транспорта а\т при биосинтезе белка, вып-ют ф-цию сигналов («запятых»), по кот-м происходит процессинг полицистронной пре-мРНК. Иногда встречаются кольцевые мультимерные (чаще всего димерные) молекулы, устроенные по типу «голова–хвост».
2) Мт‑геном высших растений представлен молекулами от 100 до 1000 Kbp.
Наличие межгенных спейсерных послед-тей. Кол.генов: 40-156.
3) Грибы и высшие растения.Присутствие помимо кольцевой молекулы (разм.мол. 20-1000 Kbp) плазмидоподобных кольцевых или лин.мол.размеры и кол.к-х может варьировать в широких пределах.
4) 6 из 10 видов пухоедов и вшей: популяция гетерогенных кольцевых молекул.
Набор миникольцевых ДНК с размером 1–2 Kbp. 1 мол.сод.1 рамку считывания. Кол.мол.к-б-ся от 100 до 1000 на кл.
5) У Кинетопластид в единственной м/х кл. им-ся сложный ассоциат: 2 популяции кольцевых молекул ДНК: миникольцевыми (размером 0,5 – 12 Kbp-105 Kbp) и максикольцевыми (20–50 молекул с размером 20–40 Kbp). Ф-ция миникольцевых мол.: кодирование малых «гидовых» РНК, которые необходимы для процесса уридилового редактирования множества первичных транскриптов максиколец.
В кл-х некот.двустворчатых моллюсков 2-х различных вариантов кольцевого мт‑генома: «мужского, M» и «женского, F», насл-е к-х происходит двумя путями: ч/з я-кл. (F) и ч/з сперму (M).
6) Лин.однокомпонентные м/х геномы, обнаруженные в ряде независимых филогенетических линий: грибы, зеленые водоросли, животные группы Cnidaria.
7) Геном - гетерогенная популяция линейных молекул. Amoebidium parasiticum, Euglenozoa (E. gracilis). 60Kbp, но выд-ся как набор коротких линейных фрагментов.
Наибольшее разнообразие хар-но для одноклеточных эукариот (Protists).
Плоидность — число одинаковых наборов хромосом, нах-ся в ядре клетки или в ядрах кл-к многоклеточного организма. Разл-т кл.гаплоидные (с одинарным набором непарных хромосом), диплоидные (с парными хромосомами), полиплоидные и анеуплоидные (когда удвоение или утрата — нулисомия — охватывает не весь геном, а лишь ограниченное число хромосом).
Транскрипция мтДНК начинается с двух промоторов тяжелой цепи (HSP1 и HSP2) и одного промотора легкой цепи (LSP). С LSP синтезируется полицистронная РНК, состоящая из восьми тРНК и одной мРНК, кодирующей субъединицу ND6, в то время как с HSP1 и HSP2 синтезируются транскрипты, включающиие остальные 14 тРНК, две рРНК и 12 мРНК, причем количество транскриптов, включающих две рРНК и две тРНК, на порядок больше. Трансляция белков, кодируемых мтДНК, происходит в матриксе на митохондриальных рибосомах (миторибосомы).
Для митохондриального генетического кода характерны те же структуры и свойства что и в случае ядерного генетического кода. Однако известны и отличия. В митохондриальной ДНК все нуклеотиды входят в состав кодонов, кодирующих либо белки, либо рРНК и тРНК. Для трансляции используется только 22 тРНК. Митохондриальная ДНК человека и других млекопитающих содержит 64 кодона, из которых 4 являются стоп-кодонами.
Каждый антикодон в случае митохондриального генетического кода способен спариваться с несколькими кодонами мРНК. Например, антикодон УАГ спаривается с кодонами ЦУУ, ЦУЦ, ЦУА и ЦУТ, кодирующими лейцин. 22 антикодона тРНК спариваются с 60 кодонами иРНК. Генетический код ДНК и белоксинтезирующий аппарат хлоропластов несколь-ко отличны от кода и белоксинтезирующего аппарата митохондрий.
Гетероплазмия.
Гетероплазмия – это присутствие в клетке различающихся молекул митохондриальной ДНК. Как правило, речь в данном случае идет об однотипных молекулах, имеющих небольшие отличия.
Различают два варианта гетероплазмии: размерную и сайтовую.
Обычно первая является результатом присутствия в контрольной области молекулы разного количества сравнительно коротких повторяющихся последовательностей (10–100 нуклеотидов). Считается, что увеличение количества повторов связано с интенсификацией процесса транскрипции мт-ДНК. Размерную гетероплазмию фиксировали у многих организмов, в частности у рыб, насекомых, летучих мышей и др.
Сайтовая гетероплазмия чаще всего связана с накоплением мутаций.
Данные об изменчивости митохондриального генома в современных этнорасовых группах человека широко используются для реконструкции истории происхождения человека. Наследование мтДНК человека строго по материнской линии и без рекомбинаций обеспечивает преемственность между поколениями и позволяет проводить генеалогический анализ популяционных данных. Получаемая информация имеет непосредственное отношение к эволюции митохондриального генофонда женской половины человечества.
Однако основанные на изменчивости мтДНК реконструкции глобальных процессов, случившихся в истории Homo sapiens, очевидно, носят универсальный характер.
Одной из важнейших проблем в происхождении рас человека является вопрос о центрах их возникновения. Существующие гипотезы первичных очагов возникновения расовых различий разделяются на моноцентрические и полицентрические в различных вариантах.
Митохондриальный геном высших растений. Особенности митохондриального генетического кода. Взаимодействие геномов митохондрий с геномом ядра. Использование митохондрий в биотехнологии.
Высшие растения имеют более крупный митохондриональный геном среди всех эукариот. У большинства высших растений и водорослей мтДНК представлена многочисленными кольцевыми молекулами размером не более 80 тыс.п.н. В митохондриальном геноме растений, помимо крупных молекул ДНК часто присутствуют кольцевые и линейные плазмиды. Большая часть генома митохондрий состоит из некодирующей последовательностей. Гены отделены друг от друга нескольким и тысячами нуклеотидных пар, поэтому каждый ген считывается автономно.
Мт‑геном высших растений представлен молекулами от 100 до 1000 Kbp. Отличия от мт жив–х касаются в первую очередь наличия межгенных спейсерных последовательностей разной протяженности и присутствием дополнительных генов. Количество идентифицированных генов колеблется от 40 до 156.
Типовой набор митохондриальных генов включает набор генов: кодирующие субъединицы комплекса I (NADH: убихинон оксидоредуктазы) nad 1–9; комплекса III (убихинон:цитохром с оксидоредуктазы) cob; комплекса IV (цитохром с оксидазы) cox 1–3; комплекса V (ATP синтазы) atp 1,6,8. Кроме того, в геноме имеются гены рРНК большой и малой субчастиц рибосом и варьирующее количество генов тРНК. Этот набор генов за редким исключением присутствует практически во всех мт‑геномах.
Митохондриальный геном растений кодирует три рибосомные РНК, 16 транспортных РНК, около 10 рибосомальных белков, некоторые белки дыхательной цепи, часть субъединиц АТФ-синтетазы, четыре белка, участвующих в синтезе цитохрома с. Наличие межгенных спейсерных последовательностей разной протяженности и присутствием дополнительных генов. Структурные гены у некоторых видов содержат интроны I или II типа. Примером этого варианта организации может быть геном диатомовой водоросли Synedra acus, геномы зеленых и красных водорослей или представителя воротничковых жгутиковых – Monosiga brevicollis.
Рекомбинации и мутации митохондриальной ДНК ведут к цитоплазматической мужской стерильности, вызывая нарушения созревания пыльцевых трубок. Интроны I, II и III группы способны к автосплайсингу и встречаются реже, чем сплайсосомные интроны. Типовой набор митохондриальных генов включает набор генов: кодирующих субъединицы комплекса I (NADH: убихинон оксидоредуктазы); комплекса III (убихинон: цитохром с оксидоредуктазы); комплекса IV (цитохром с оксидазы); комплекса V (ATP синтазы). В геноме имеются гены рРНК большой и малой субчастиц рибосом и варьирующее количество генов тРНК. Среди дополнительных генов следует отметить: от 3 до 27 генов рибосомных белков (чаще малой субчастицы), гены субъединицы комплекса II (сукцинат:убихинон оксидоредуктазы), уникальные неидентифицированные рамки считывания.
В том случае, если все митохондрии в клетке имеют одинаковую копию ДНК дикого типа – наблюдается гомоплазмия (однотипность митохондриальной ДНК у индивидов). Гетероплазмия – это присутствие в клетке различающихся молекул митохондриальной ДНК. Различают два варианта гетероплазмии: размерную и сайтовую.
Все гены процессинга РНК, транскрипции, трансляции перенесены в ядро.
Транскрипция мтДНК начинается с двух промоторов тяжелой цепи (HSP1 и HSP2) и одного промотора легкой цепи (LSP). С LSP синтезируется полицистронная РНК, состоящая из восьми тРНК и одной мРНК, кодирующей субъединицу ND6, в то время как с HSP1 и HSP2 синтезируются транскрипты, включающиие остальные 14 тРНК, две рРНК и 12 мРНК, причем количество транскриптов, включающих две рРНК и две тРНК, на порядок больше. Трансляция белков, кодируемых мтДНК, происходит в матриксе на митохондриальных рибосомах (миторибосомы).
Для митохондриального генетического кода характерны те же структуры и свойства что и в случае ядер-ного генетического кода. Однако известны и отличия. В митохондриальной ДНК все нуклеотиды входят в состав кодонов, кодирующих либо белки, либо рРНК и тРНК. Для трансляции используется только 22 тРНК. Митохондриальная ДНК человека и других млекопитающих со-держит 64 кодона, из которых 4 являются стоп-кодонами.
Каждый антикодон в случае митохондриального генетического кода способен спариваться с несколькими кодонами мРНК. Например, антикодон УАГ спаривается с кодонами ЦУУ, ЦУЦ, ЦУА и ЦУТ, кодирующими лейцин. 22 антикодона тРНК спариваются с 60 кодонами иРНК. Генетический код ДНК и белоксинтезирующий аппарат хлоропластов несколько отличны от кода и белоксинтезирующего аппарата митохондрий.
Экспрессией большинства генов митохондрий управляют определенные гены ядер. Если блокировать синтез белка в митохондриях интактных клеток, то в цитоплазме начинают в избытке образовываться ферменты участвующие в митохондриальном синтезе ДНК, РНК и белков, клетка пытается преодолеть воздействие блокирующего агента. РНК редактирование в митохондриях цветковых растений заключается в дезаминировании С до U. Дезаминирование осуществляется специализированным ферментом класса дезаминаз. Реакция дезаминирования C→U происходит строго сайт-специфично. Репликация осуществляется с участием белков, кодируемых яДНК, – митохондриальной ДНК-полимеразой, геликазой и белком, связывающимся с оцДНК.