|
|
РЕЦЕПТОРЫ СТВОЛА МОЗГА
Сто лет назад Клод Бернар (1865) положил начало новой ветви нейрофизиологии, связанной с изучением регуляции метаболизма и эндокринных функций организма со стороны нервной системы. Эти знаменитые теперь «острые опыты», в которых диабет вызывался уколами в ствол мозга, привели к множеству других экспериментов, в которых нарушения вегетативных функций тела и внутренней среды были следствием небольших повреждений в стволе мозга или его локальной электрической стимуляции (рис. IX-3).
Так, наблюдались изменения температуры тела, осмотического давления тканевой жидкости, функции гипофиза и других эндокринных желез, а также изменения уровня сахара в крови. Когда постепенно была изучена организация этих нейронных систем, стало очевидным, что группы клеток в стволе мозга действуют как рецепторные зоны в отношении субстанций или переменных, которые они контролируют. Так было обнаружено, что инъекции гипертонического раствора соли в третий желудочек вызывает у козы обильное поглощение воды (Andersson, 1953). Тепло, приложенное к основанию переднего края третьего желудочка, сразу же ведет к изменению в механизмах терморегуляции всего тела млекопитающего (Ranson, Fisher and Ingram, 1937); инъекции андрогенов и эстрогенов, меченных радиоактивными молекулами, непосредственно позади этой области сопровождаются различным их поглощением клетками мозга, стимуляция которых вызывает половое поведение (Michael, 1962; Davidson, Jones and Levine, 1968; см. также рис. 1Х-4); сходным образом меченые молекулы глюкозы максимально усваиваются труппой клеток, расположенных еще дальше назад и контролирующих процесс поглощения пищи (см. гл. X; Mayer, 1963), а локальные изменения в парциальном давлении СОг в задней части ствола мозга вызывают резкие изменения частоты и глубины дыхания (Meyer, 1957). Все эти виды чувствительности локализованы в структурах, которые расположены в стволе мозга, довольно близко к желудочкам средней линии (третьему ж четвертому).
Тот факт, что эта часть мозга содержит рецепторы, не должен казаться столь удивительным. В ходе эмбриогенеза эта часть центральной нервной системы, расположенная по средней линии, развивается из наружного зародышевого листка, из которого формируется вся центральная нервная система Эта ткань имеет такое же происхождение, как и кожа: гребешок экто дерма льных клеток на спине эмбриона складывается таким образом, что образует трубку, полость которой позднее будет заполнена цереброспинальной жидкостью (рис. IX-5). В головном конце эмбриона эта полость становится системой желудочков мозга. Следовательно,

Рис IX-З. Схема ствола мозга на фоне контура полушарий головного мозга (Livingston, 1955).
стенки полости, перивентрикулярные клетки сродни эктодерме, из которой образуется кожа и некоторые более специализированные рецепторы, такие, как сетчатка. Таким образом, чувствительность перивентрикулярных структур сходна с чувствительностью кожи; температурные изменения, деформация, изменения водного обмена — некоторые из главных категорий стимулов, к которым дувствительны и перивентрикулярные структуры и кожа. Об этой чувствительности перивентрикулярной части мозга мне стало известно в результате одного драматического происшествия. Одной из особенностей мозговой ткани является то, что она почти в любом месте нечувствительна к механическим воздействиям. Поэтому операция на мозге в большинстве случаев делается под местной анестезией, чтобы уберечь пациента от большой травмы и от риска, связанного с общей анестезией. В одной из таких операций мы исследовали область четвертого желудочка и, как обычно при нейрохирургической операции, сохраняли мозг влажным с помощью жидкости, капавшей на его обнаженные части. Раствор, который обычно используется, имитирует концентрацию и химический состав цереброспинальной жидкости; в данном случае неопытная медицинская сестра случайно заменила его дистиллированной водой. В тот момент, когда

Рис. 1Х-4. Иллюстрация расположения рецепторов в стволе мозга крысы: точками показаны области, содержащие радиоактивно меченные молекулы эстрогена (женский половой гормон). Латерально расположенная структура с высокой концентрацией эстрогена — амигдала (Stumpf, 1970).
вода достигла желудочка, пациент испытал резкую боль в голове, тошноту, позывы к рвоте и рвоту. Такие же сильные реакции вызывались легким вдавливанием или вытягиванием стенки желудочка или использованием жидкости, которая была холоднее или теплее температуры тела.
Таким образом, мы можем сделать вывод, что сотни нейрофизиологических экспериментов показывают, что по средней линии ствола мозга близко к системе желудочков расположены группы
Рис, IX-5, Схема формирования нервной трубки, из которой в дальнейшем развивается спинной и головной мозг (Truex, Carpenter, 1969).

специализированных «контролирующих» рецепторов. Эти специализированные рецепторы являются классическими центрами контроля дыхания, насыщения и т. д., то есть всего того, что интересует психологов и биохимиков, изучающих регуляцию метаболизма и эндокринных функций организма со стороны нервной системы. Эти рецепторы функционируют как чувствительные к «состоянию» организма элементы класса сервомеханизмов, названных Кенноном (1929) «гомеостатами», которые имеют отношение к регуляции функций, включающих подготовительный и завершающий этапы. Теперь мы перейдем к рассмотрению некоторых других компонентов гомеостатов.
КОРРЕКЦИЯ ГОМЕОСТАТИЧЕСКИХ МЕХАНИЗМОВ
Непосредственно за пределами перивентрикулярных рецепторов находится сплетение нервных волокон. В него в виде пятен вкраплены группы нервных клеток, и его пересекают длинные нервные волокна. Анатомия ретикулярной формации среднего-мозга была детально рассмотрена А. Бродалем (1958) и Шейбе-лами (1958); ее физиология подробно обсуждалась Джаспером (1958) и Мэгуном (1965). Однако часто упускается из виду тот факт, что организация, подобная той, которая была найдена в среднем мозге, тянется далеко вперед от него в передний мозг вдоль желудочков средней линии. Таким образом, части гипоталамуса и таламуса, расположенные по средней линии, и даже область перегородки, обладают такими же свойствами, как ретикулярная формация среднего мозга. В конечном счете разделение мозга на задний, средний и передний весьма условно. Столь же возможной, а для проблемы гомеостатических регуляций даже более важной является классификация, согласно которой мозг делится на пласты в направлении от внутренней части к наружной, аналогично пластам в удлиненной луковице. Характерно, что ткань ствола мозга состоит из нейронов с весьма короткими и тонкими волокнами, которые имеют обширные дендритные сети (рис. IX-6). На каждой нервной клетке сходятся многие ответвления от классических длинных проекционных путей, берущих начало от различных сенсорных рецепторов организма. Как было показано в экспериментах с регистрацией нейронной активности посредством микроэлектродов, каждый нервный элемент системы находится под влиянием многих сенсорных модальностей (рис. IX-7). Кроме того, существуют реципрокные отношения между глубокими структурами мозга и остальной частью нервного сгвола; например, кора головного мозга активируется при электрическом раздражении глубоких отделов мозга, и, наоборот, стимуляция коры изменяет активность ретикулярных систем. Такая конвергенция сенсорных влияний, а также диффузный характер»

Рис. IX-6. Поперечный срез продолговатого мозга (выше его трети) у 10-дневного котенка. На нем видны конвергенция и перекрытие окончаний афферентных волокон в ретикулярной формации ствола мозга. Слева — небольшая группа волокон зарисована непосредственно под микроскопом, справа —схема перекрывающих друг друга волокон (Scheibel and Scheibel, 1967).
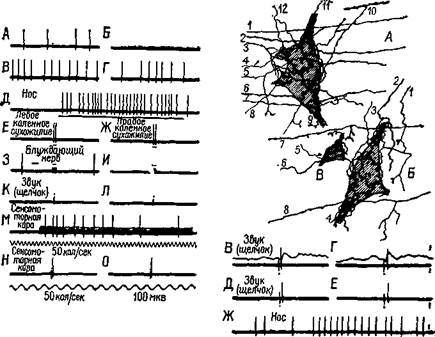
Рис. 1Х-7. Физиологическая и гистологическая демонстрация конвергенции гетерогенных афферентов на одиночные нейроны ретикулярной формации ствола мозга. Фрагменты записи от А до О и от В до Ж иллюстрируют динамические структуры экстраклеточных спаиковых разрядов двух нейронов в ретикулярной формации продолговатого мозга. А — спонтанные разряды; Б — торможение, вызванное поляризацией головного мозга (передняя доля, поверхность позитивна); В — эффект «отдачи», следующий за прекращением поляризации; Г — возвращение к нормальной спонтанной активности; Д — реакция, вызванная давлением на нос; Е и Ж — реакция на двусторонние удары по коленному сухожилию; 3 и И — отсутствие эффекта на короткую стимуляцию блуждающего нерва; К и Л — отсутствие реакции на щелчки; М — реакция вызывается повторной кортикальной стимуляцией; Н и О — реакция с коротким латентным периодом на одиночный удар электрического тока, приложенного к коре (обратите внимание на разный масштаб времени для этих двух записей). Записи В, Г, Д н Е (справа) относятся к другой клетке, которая чувствительна к давлению на нос, а также к щелчкам. Нейроны А, Б и В — нейроны продолговатого мозга 10-дневного котенка. На рисунке частично показаны синапсы окончаний афферентных волокон. Горизонтально ориентированные волокна, такие, как 1—7 (А). ш. 6—8 (Б), могут принадлежать к длинным спинноретикулярным и ретикуло-ретикулярным волокнам, тогда как волокна типа 1—4 (Б) являются сенсорными коллатералями и окончаниями волокон, идущих из мозжечка (Scheibel and Scheibel, 1967).
взаимодействия говорят о том, что наиболее вероятной функцией этих систем является регуляция общего состояния возбудимости нервной системы. Это предположение подтверждается данными о том, что электрическая стимуляция систем глубоких отделов мозга вызывает активацию и дезактивацию ритмов коры, а также тем, что была обнаружена связь разрушения и стимуляции этих систем с такими психологическими процессами, как цикл сон — бодрствование и тревога. Более того, анатомическая структура этих систем свидетельствует о том, что механизмы градуального ответа, характеризующие микроструктуру медленных потенциалов, доминируют в них над механизмами проведения сигналов. Они обильно насыщены синапсами и дендритами, включают в основном короткие и тонкие волокна, поэтому скорость проведения импульса в них невелика, а его амплитуда мала. Как было отмечено в первой части книги, микроструктура медленных потенциалов особенно чувствительна к изменениям в химической среде. Как мы вскоре увидим, большое число работ посвящено изучению действия нервных трансмиттеров и психофармакологических веществ на функции этих систем. Но сначала рассмотрим, какое значение имеет ретикулярная формация из-за ее близости к специализированным перивентрикулярным рецепторам для проблемы гомеостатической регуляции внутренней среды организма.
Внутри мозгового ствола расположен ряд систем, которые играют особую роль в регуляции гомеостатического механизма. История этого важного открытия даст нам некоторое представление о том, как какое-то неожиданное событие может повлечь за собой фантастические научные открытия.
Двое исследователей Университета Мак-Гилла, Дж. Олдс и П. Милнер (1954), готовились провести электрическую стимуляцию ретикулярной формации ствола 'мозга крыс во время обучения этих животных решению задач. С помощью стереотаксиче-•ского прибора исследователи вживили электроды в те зоны мозга крыс, которые, по их мнению, были наиболее подходящими для опыта. В предварительных опытах, изучая поведение крыс, исследователи заметили, что при включении электрического импульса крыса всякий раз убегает в определенное место. Это озадачило и заинтересовало исследователей. После многократного наблюдения такого эффекта им захотелось автоматизировать методику с тем, чтобы на досуге подробно изучить это «принудительное повторение». Они изменили ситуацию таким образом, что крыса могла обнаружить педаль в углу, нажатие на которую включало импульс тока, раздражающего мозг. Крыса быстро научилась находить педаль и нажимать на нее. Таким образом появилась на свет методика самораздражения мозга.
Затем Олдс с большой тщательностью составил карты участков мозга, от которых мог быть получен этот эффект. Но

Рис. 1Х-8. Медиальный (верхняя схема) и латеральный (нижняя) сагиттальные срезы мозга крысы, показывающие локализацию центров положительного подкрепления в опытах с самораздражением. Квадратиками и кружками обозначен процент двигательных реакций (нажатия на педаль) в течение 6-часового эксперимента. А — передний таламус; AM — амигдала; С — хвостатое ядро; СВ — мозжечок; СС — мозолистое тело; С CORT — цин-гулярная кора; Е — энториальная кора, FX — свод; НРС — гиппокамп^ НТН — гипоталамус; MB — мамилярное тело; МТ — мамилоталамический тракт; PREPYR — препириформная кора; S — перегородка; TEG — тегмен-тум; ТН — таламус. Медиальный срез (верхний) сделан вблизи средней линии, латеральный (нижний) — на 2—3 мм в сторону от нее. Буквы М (медиально) и L — (латерально) около других обозначений указывают на то, что структура находится примерно на 2 мм медиапьнее или лате-ральнее по отношению к представленному срезу (Olds, 1961).
стереотаксическая техника не дала ожидаемых результатов,— место расположения стимулирующего электрода оказалось далеко впереди от намеченного. Но участок мозга, так случайно найденный — медиальный переднемозговой пучок в области перегородки, — оставался одной из главных зон для получения этого эффекта. Остальная часть системы самораздражения простирается назад от этой зоны и включает, по-видимому, участки ствола мозга с рецепторными механизмами, уже подробно рассмотренными в этой главе (Olds, 1961, рис. IX-8).
Было проведено много экспериментов с использованием техники самораздражения, и было дано много толкований этого эффекта, начиная от простых гедонистических утверждений, что открыты «центры удовольствия», до осторожных бихевиористских предположений о подкрепляющих свойствах этого процесса. Моя собственная точка зрения частично основывается на сообщениях, полученных от наблюдений над больными с вживленными в эти участки мозга электродами, а также на анатомических данных и результатах исследования поведения, говорящих о том, что локализации электродов для эффективной самостимуляции и для контроля за таким инстинктивным поведением, как, например, пищевое, по существу, совпадают. Исходя из этих данных, я рассматриваю самораздражение мозга как мгновенное изменение системы регуляции, как такую коррекцию базальных гомеостатических механизмов, в результате которой организм, ощущая временный голод, жажду и т. п., затем быстро испытывает мгновенное насыщение только для того, чтобы повторить цикл еще раз. Специфика ощущений, вызываемых стимуляцией, зависит от рецепторной системы ствола мозга, примыкающей к участку, выбранному для самораздражения, и она приводит к соответствующей деятельности, если для этого имеются возможности (Olds, 1955). Следовательно, процесс самораздражения напоминает регулирующее устройство в домашнем термостате, которое повторно устанавливается и возвращается в исходное положение в комнате, где уже стало тепло. Топка включается лишь на короткое время, чтобы затем снова выключиться, как только показатель на регуляторе достигнет своего исходного значения.
Коррекция гомеостатических механизмов с помощью электрической стимуляции не является единственной. Как уже отмечалось, в ряде работ инстинктивное поведение вызывалось также небольшими инъекциями химических веществ. Участки, реагирующие на химическую стимуляцию, и тут совпадали с участками, с которых получали эффект электрического самораздражения. Побочно наблюдавшиеся специфические эффекты зависели от близости стимулируемого участка к той или другой рецепторной системе ствола мозга. Эти эксперименты показывают, что могут быть приведены в действие по крайней мере два различных
корригирующих процесса: один из них имеет андренэргическую природу и сконцентрирован главным образом в среднем мозге; а другой — холинэргическую и концентрируется в более передних частях ствола мозга.