|
|
У корі людини налічується приблизно 600 млн таких колонок.
Наявність колонок, пристосованих для сприймання інформації, не заперечує існування у корі інших систем, які виконують ефекторні функції. До них належать системи, що оперують різними медіаторами (адренергічна, холінергічна, дофамінергічна тощо).
Ділянки кори, маючи шестишарову будову, відрізняються за своєю цитоархітектонічною картиною.
За цим показником уся кора великого мозку людини поділена на 52 окремі ділянки — поля (мал. 170). Причому різні за будовою поля виконують різні функції.

Розглянемо деякі з найважливіших ділянок нової кори, які входять до основних часток.
У корі лобової частки півкуль великого мозку розміщені:
- передцентральна (рухова — моторна — і премоторна),
- власне лобова (префронтальна) ділянки, пов'язані зі специфічно людською мовною функцією.
У задньому відділі нижньої лобової звивини розміщена зона Брока — руховий центр мови, який разом із центром Верніке (у 95 % людей вони розміщені у лівій півкулі) забезпечують здатність людини читати, писати, чути, вимовляти і розуміти мову.
Кора лобової частки відіграє вирішальну роль у набуванні нових навичок, управлінні поведінкою в нестандартних ситуаціях. В основі такої функції лежить здатність нейронів кори адекватно реагувати на упорядковані послідовності подій.
У корі лобової частки виділяють десять цитоархітектонічних полів з нечіткою межею. Довгі асоціативні волокна сполучають лобову ділянку з усіма іншими відділами кори великого мозку, таламусом, гіпоталамусом, хвостатим ядром, структурами лімбічної системи. Ця величезна система зв'язків забезпечує можливість інтеграції інформації із зовнішнього та внутрішнього середовищ.
Ушкодження різних полів кори лобової частки спричинює різні ефекти. У людей у випадках патології кори лобової частки найчастіше спостерігаються втрата ініціативи, апатія, порушення абстрактного мислення, нездатність до творчого мислення, стереотипність у поведінці й висловлюваннях, розгальмовування нижчих емоцій і потягів, некритичне ставлення до своїх вчинків, розлади мовлення і понятійного мислення, значні зміни особистості.
Соматосенсорна кора — це вищі (кіркові) рівні рухового і шкірного аналізаторів. Вони утворюють цілісну структурно-функціональну систему, за допомогою якої контролюється і регулюється активність і аналізується вся інформація, що надходить від будь-якої точки тіла.
У корі за центральної звивини тім'яної частки розміщена перша соматосенсорна зона, куди через специфічні ядра таламуса потрапляють волокна висхідних шляхів спинного мозку. Тут знаходяться центри шкірної і пропріоцептивної чутливості.
Рухова (моторна) зона кори розміщена у передцентральній звивині, яка лежить попереду від центральної (роландової) борозни.
Рухова кора відрізняється від інших її відділів передусім своєю товщиною, яка досягає 3-4,5 мм і в якій знаходяться великі пірамідні клітини (Беца) (до 1000 мкм діаметром), що містяться у V пластинці (шарі). Великі й дрібні пірамідні клітини у III пластинці (шарі) кори дають початок кірково-спинномозковим шляхам, їхні аксони прямують до внутрішньої капсули, а дендрити піднімаються до поверхні кори. Гігантські пірамідні клітини проводять збудження через кірково-сиинномозкові шляхи зі швидкістю 60-90 м/с, але вони становлять лише З % загальної кількості волокон цих шляхів у кожній півкулі. Решта волокон проводить збудження значно повільніше.
Існує упорядкована просторова проекція різних частин тіла на рухову ділянку кори людини, причому зони, які керують рухами руки, мускулатури лиця, губ і язика, займають найбільшу поверхню кори ( руховий гомункулюс).
Отже, в корі обох півкуль існують чотири чутливі та чотири рухові зони.
У корі потиличної частки розміщений кірковий кінець зорового аналізатора, центральна частина якого у людини локалізується в ділянці шпорної борозни.
Вторинна і третинна зорові зони оточують первинну зону кільцем. Особливістю будови первинної зорової зони є значний розвиток IV шару кори, який складається з численних зірчастих нейронів, і розщеплення цього шару на три підшари.
У первинній проекційній зоні закінчуються зорові волокна, що перериваються у бічному колінчастому тілі, а у вторинній зоні — волокна, які перериваються в ядрах подушки таламуса. Ці дві системи зорових проекцій значною мірою ізольовані одна від одної.
Первинна зорова зона потрібна для чіткого сприймання зорових об'єктів, а вторинна здійснює впізнання зорових образів, тут зберігається зорова пам'ять. У третинній зоровій зоні (19 поле) відбувається впізнання символічних зорових образів (літер, цифр, міміки обличчя тощо).
У корі великих півкуль за функціональними ознаками розрізняють проекційні й асоціативні поля.
Проекційні поля
становлять центральні (кіркові) кінці аналізаторів.
Це:
- І і II сенсомоторні зони;
- мотосенсорні зони ;
- зорова зона,
- слухова зона;
- смакова зона,
- нюхова зона кори займає ділянку морського коника і прилеглих ділянок кори.
Всю іншу поверхню кори займають асоціативні поля.
Асоціативні поля
— ділянки кори, не пов'язані з виконанням якоїсь специфічної сенсорної чи рухової функції.
Асоціативні поля кори великого мозку займають більшу площу, ніж проекційні.
Нейрони цих полів беруть участь в інтеграції сенсорної інформації та забезпеченні зв'язків між чутливими й руховими зонами кори.
Функція асоціативних полів є набагато складнішою.
Їхні нейрони, отримуючи інформацію від різних аферентних систем, об'єднують її, інтегрують, формують і запам'ятовують узагальнюючі, абстрактні поняття та уявлення.
Асоціативні поля значною мірою визначають поведінку людини, її ставлення до навколишнього середовища, до інших людей, інтелектуальний і культурний рівень особистості.
Кора тім'яної частки майже вся зайнята асоціативними полями. У разі ушкодження цієї ділянки страждають найскладніші процеси аналізу й синтезу вищої нервової діяльності.
Тім'яна частка — це апарат вищої інтегративної діяльності мозку людини, вона безпосередньо стосується процесів біологічної й соціальної адаптації, є фізіологічною основою вищих психічних функцій.
Електрична активність головного мозку.
Запис сумарної електричної активності головного мозку, яка відводиться за допомогою зовнішніх електродів, називають електроенцефалограмою (ЕЕГ). У ній розрізняють кілька основних ритмів:
альфа-ритм (8-12 Гц), який реєструється у стані спокою при заплющених очах;
бета-ритм (13-30 Гц) — у стані спокою з розплющеними очима;
гамма-ритм (30-70 Гц) — під час напруженої розумової роботи;
тета-ритм (4-7 Гц) — у напруженому емоційному стані;
дельта-ритм (0,5-3 Гц) — під час глибокого сну (мал. 172, а).
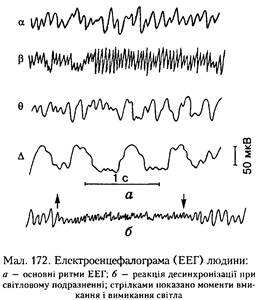
Коли на ЕЕГ помітні переважно альфа-або дельта-ритми, це явище називають синхронізацією, а бета- і гамма-ритми — десинхронізацією електричної активності мозку (див. мал. 172, б).
ЕЕГ відображає переважно повільні зміни МП кіркових нейронів, особливо ЗПСП та ГПСП. Оскільки реєструвальні електроди розміщені на значній відстані від джерела електричних потенціалів, тобто клітинних мембран, то амплітуда коливань ЕЕГ у 100-1000 разів менша, ніж значення МП, яке реєструється внутрішньоклітинно.
ЕЕГ відображає сумарні потенціали, які відводяться одночасно від значної кількості нейронів. Так, електрод діаметром близько 1 мм реєструє активність близько 100 000 нейронів, розміщених на глибині 0,5 мм.
Багато дослідників вважають, що альфа-ритм виникає спочатку в таламусі, а звідти поширюється на кору, оскільки руйнування таламо-кіркових шляхів чи видалення таламуса призводять до зникнення альфа-хвиль в корі півкуль головного мозку. У той же час ритмічна активність нейронів таламуса зберігається після декортикації. Проте деякі дослідники наполягають па тому, що ритмічна електрична активність зумовлена функцією самих нейронів кори великого мозку.
Основні параметри ЕЕГ, зокрема альфа-ритм, мають чіткі індивідуальні ознаки, що дає змогу віднести окрему людину до певного типу за особливостями її ЕЕГ.
Окремі ритми ЕЕГ пов'язані з проявом різних видів розумової діяльності людини.
Так, посилення тета-ритму пов'язане з емоційною напруженістю суб'єкта, зокрема з відчуттям задоволення чи болю. У звичайних умовах у дорослих людей з урівноваженим характером тета-ритм мало помітний, але під час дії неприємних подразників цей ритм чітко виявляється навіть у стриманих людей.
У мозку людини виділяють також повільні ритмічні коливання хвилинного діапазону — надповільні електричні потенціали:
дзета-хвилі (5-12 за 1 хв),
тау-хвилі (14 за 1 хв) та
іпсилон-хвилі (0,5-4 за 1 хв).
Деякі з цих потенціалів зростають під час напруженої розумової діяльності, деякі реєструються під час сну, а також у гіпнотичному стані (стадія сомнамбулізму).
Плоска, або нульова, ЕЕГ є критерієм смерті мозку. Така ЕЕГ реєструється також після застосування електрокопвульсивного шоку (до ЗО хв).
Функціонально-структурна організація мозочка, його аферентні та еферентні зв’язки, їхня фізіологічна роль. Роль мозочка в програмуванні, ініціації та контролюванні рухів. Наслідки видалення або ураження мозочка.
Мозочок є типовою надсегментарною структурою, аферентні та еферентні зв'язки якої починаються і закінчуються в інших утворах мозку. Він здійснює регуляцію рухових функцій.
У зв'язку з вертикальним положенням тіла у людини розміри і значення мозочка істотно зростають.
Мозочок міститься під потиличними частками півкуль великого мозку і лежить у задній черепній ямці. У ньому розрізняють дві півкулі і черв'як, їх поверхні вкриті поперечними борознами, між якими розташовані довгі звивини мозочка.