|
|
Генез ,особенности структуры , свойства и функции тканевых базофилов
Тучные клетки (мастоциты) и базофилы представляют тканевые клетки, содержащие в цитоплазме базофильные гранулы. Оба типа клеток имеют костномозговое происхождение и принадлежат к миелоидному ряду.. В отличие от базофилов, относящихся к клеткам крови, тучные клетки не циркулируют в крови и представляют тканевые клетки. Мастоциты реагируют на разного рода повреждающие воздействия, участвуют в развитии воспаления, служат основными эффекторными клетками при гиперчувствительности немедленного типа и входят в первую линию иммунной защиты, обеспечивая в первую очередь защиту от многоклеточных паразитов. Базофилы также могут выполнять аналогичные функции. Однако если тучные клетки, находясь в очагах повреждения, реагируют на него немедленно, вовлечение базофилов в подобные реакции требует их миграции в ткани, что исключает участие базофилов в инициации и осуществлении ранних этапов реакций врожденного иммунитета. Предполагают, что у тучных клеток и базофилов есть общий предшественник. Однако неясно, развивается ли он непосредственно из общего миелоидного предшественника или служит ответвлением одного из основных направлений миелоидной дифференцировки (эозинофильно-базофильного). Схема развития тучных клеток и базофилов представлена на рис. 2.4.Согласно этой схеме окончательная дифференциация предшественников этих клеток происходит в селезенке. Базофилы могут созревать как в костном мозгу, так и в селезенке, и мигрируют в кровоток. Дифференцировка тучных клеток проходит иначе: в кровоток поступают предшественники тучных клеток (у человека эти клетки в циркуляции имеют фенотип CD13+CD33+ CD34+ CD38+ CD117+). Из кровотока предшественники тучных клеток мигрируют в ткани (в наибольшем количестве — в слизистую оболочку кишечника), где и завершается созревание мастоцитов. Основные факторы, определяющие дифференцировку тучных клеток — SCF и IL-3; в качестве кофакторов выступают IL-4, IL-9, IL-10 и фактор роста нервов (NGF). В частности, эти факторы обусловливают формирование гранул и пролиферацию клеток. В слизистых оболочках в роли фактора, необходимого для развития тучных клеток, выступает IL-33. Тучные клетки сохраняют способность к делению и имеют длительный срок жизни — месяцы и даже годы. Диаметр тучных клеток варьирует от 10 до 20 мкм. Они имеют овальную форму с ворсинчатой поверхностью. Мембранный фенотип тучных клеток выражается формулой FcεRI+ CD13+ CD29+ CD45+ CD117+ СD123+. Среди мембранных молекул тучных клеток наиболее важны для реализации их функции высокоаффинные рецепторы IgE — FcεRI. Как уже было сказано, главная морфологическая особенность этих клеток — наличие в их цитоплазме большого количества базофильных гранул(10–150 на клетку). Гранулы разновидностей тучных клеток варьируют по составу , однако они всегда содержат вазоактивные амины, главный из которых — гистамин — реализует значительную часть эффектов тучных клеток при аллергических реакциях. Кроме того, в гранулах содержатся хондроитинсульфаты А и С и/или гепарин, а у некоторых видов животных (например, у кроликов) — серотонин. В состав гранул входят также ферменты: прежде всего протеазы, а также дегидрогеназа, пероксидаза, РНКаза, гистидинкарбоксилаза и кислые гликозамингликаны. Выделяют 3 группы протеаз тучных клеток: триптазы (ферменты со специфичностью, близкой к трипсину; у мышей — 5 разновидностей), химазы (сходны по специфичности с химотрипсином; у мышей — 4 варианта) и карбоксипептидазу А (относится к металлопротеиназам). Перечисленные факторы,содержащиеся в гранулах, — предобразованные вещества. Перекрестное cвязывание рецепторов FcεRI комплексами IgE-антител с аллергенами обусловливает высвобождение содержимого гранул (дегрануляцию) и проявление всех основных реакций гиперчувствительности немедленного типа. Дегрануляция может быть вызвана также повышением__содержания внутриклеточного цАМФ или концентрацией в цитозоле ионов Са2+. Дегрануляция не сопровождается гибелью клеток — гранулы после выброса регенерируют. Тучные клетки несут некоторые патогенраспознающие рецепторы (TLR-2, TLR-3, TLR-4), что позволяет им распознавать патогены и их продукты напрямую. При стимуляции тучные клетки синтезируют и секретируют эйкозаноиды и цитокины. Из эйкозаноидов в тучных клетках в наибольшем количестве вырабатываются лейкотриен С4 и простагландин E2. Спектр цитокинов, секретируемых тучными клетками, сходен со спектром цитокинов, продуцируемых Т-хелперами 2-го типа : IL-3, IL-4, IL-5, IL-6, IL-10, IL-13, GM-CSF. Тучные клетки вырабатывают также провоспалительные (IL-1, IL-8, IL-12, IL-18, IL-21, IL-23, TNFα) и гомеостатические цитокины (IL-7 и IL-15), а также TGFβ, некоторые хемокины и интерфероны основных типов. IL-4, TNF и GM-CSF мастоциты вырабатывают спонтанно, образование остальных цитокинов индуцируется стимуляторами. Активированные тучные клетки продуцируют ряд пептидных ростовых факторов (сосудистый — VEGF, фибробластный — FGF, фактор роста нервов — NGF). Спектр секертируемых цитокинов (особенно спонтанная выработка IL-4) определяет иммунорегуляторную функцию тучных клеток, главное проявление которой — участие в индукции дифференцировки Th2-клеток.Для тучных клеток характерны поверхностные маркеры: CD117 (с-Kit) —рецептор для SCF и CD123 — рецептор для IL-3. SCF и IL-3 (помимо их роли в качестве факторов, определяющих развитие тучных клеток) служат основными факторами роста зрелых мастоцитов. Тучные клетки несут на своей поверхности также высокоаффинные FcγI-рецепторы и рецепторы для компонентов комплемента C3b и С3d (мукозные тучные клетки лишены CR1), что свидетельствует об их участии в реакциях врожденного иммунитета. На поверхности тучных клеток присутствуют молекулы МНС обоих классов; наличие МНС-II, а также костимулирующих молекул CD86 придает мастоцитам способность выполнять функции АПК, особенно при индукции Th2-клеток. Тучные клетки локализуются в подслизистом слое слизистых оболочек (особенно в кишечнике), соединительнотканном слое кожи (дерме), серозных оболочках, селезенке, периваскулярной соединительной ткани. В 1 г названных тканей содержится 104–106 тучных клеток. Мастоциты легко идентифицировать по окрашиваемости толуидиновым синим или алциановым синим. Выделяют два варианта тучных клеток: слизистые, или мукозные (тип t), и серозные (тип сt) (табл. 2.4). Названия отражают 2 главных отличительных признака этих клеток — преимущественную локализацию и преобладающий тип протеаз (триптазы — t или хемотриптазы — ct). Оба типа тучных клеток происходят из костного мозга, но только клетки t-типа в своем развитии зависят от тимуса и отсутствуют у генетически бестимусных мышей. Продолжительность жизни серозных тучных клеток выше, чем слизистых. Основной ростовый фактор для клеток обоих типов — SCF; в качестве кофактора для слизистых тучных клеток выступают IL-3 и IL-4, для серозных — только IL-3. Преобладающий тип протеогликана в слизистых тучных клетках — хондроитинсульфат,в серозных — гепарин. На поверхности мукозных мастоцитов экспрессировано больше FcεRI, они содержат больше IgE в цитоплазме, чем__серозные. Тучные клетки разных типов различаются также интенсивностью секреции эйкозаноидов: в слизистых тучных клетках больше лейкотриенов, в серозных — простагландина. Несмотря на существенные различия, до конца не известно, являются ли эти разновидности тучных клеток истинными субпопуляциями или представляют фенотипические варианты единой популяции тучных клеток, дифференцирующиеся под влиянием факторов микроокружения. У разных типов тучных клеток микроокружение различается: мастоциты типа t локализованы главным
образом в подслизистом слое мукозы, а тучные клетки типа ct — в серозных полостях, дерме и миндалинах. Участие в защите от паразитов и развитии аллергических реакций доказано только для слизистых тучных клеток (типа t), тогда как серозные мастоциты причастны скорее к развитию склеротических процессов.
30.
Лимфоциты – небольшие мононуклеарные клетки, координирующие и осуществляющие иммунный ответ за счет продуцирования воспалительных цитокинов и антигенспецифических связывающих рецепторов. Одной из групп лимфоцитов являются В-л. В-л. и их наиболее зрелые формы - плазматические клетки - продуцируют иммуноглобулины (антитела), то есть осуществляют синтез
эффекторов гуморального иммунитета. Специфическими рецепторами В-лимфоцитов являются молекулы иммуноглобулинов. На В-лимфоцитах lg-рецепторы нековалентно ассоциированы с двумя трансмембранными белками - Iga и Igp или Iga и Igy. Молекулы lg и полипептидные цепи,входящие в состав В-клеточного рецептора, являются наиболее надежными маркерами В-линейной принадлежности.


В-л. Осуществляют экспрессию уникальных антигенных рецепторов – иммуноглобулинов – и запрограммированы на продукцию их в большом количестве в ответ на антигенную стимуляцию. В-л. Образуются из стволовых клеток костного мозга, созревание В-д. у человека происходит в основном в костном мозге. ИС содержит большую популяцию отдельных клонов В-л., каждый из которых экспрессирует уникальный антигенный рецептор. Разнообразие клонов В-л. Обеспечивает разнообразие вырабатываемых ими антител.
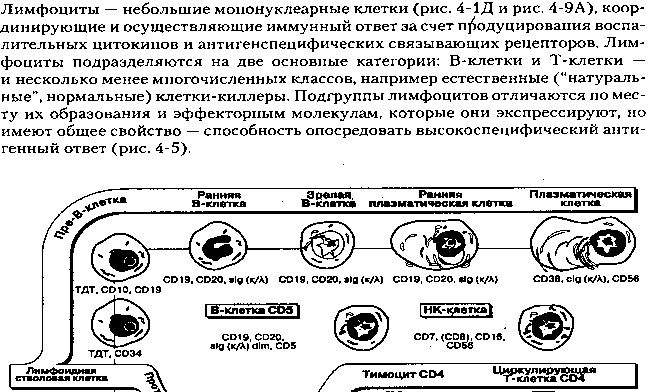
Дифференцировка.B-клетки все стадии антигеннезависимой дифференцировки проходят в костном мозге . На поверхности предшественников B-лимфоцитов, про-В- лимфоцитов , обнаруживают ряд CD , однако данные об их экспрессии противоречивы. Наиболее ранние про-В-клетки часто определяют как CD19плюсCD10плюс-клетки, не экспрессирующие генов тяжелых цепей иммуноглобулинов, но экспрессирующие антигены MHC класса II . Возможными кандидатами для определения про-В-клеток являются CD9 , а также CD24 : экспрессия CD24 (как и CD10 ) не ограничена клетками B-ряда, но ее уровень на ранних этапах дифференцировки повышен. CD19 является наиболее универсальным маркером клеток B-лимфоцитарного ряда (так называемый пан- B ) - он обнаруживается уже на поверхности B-клеток эмбриональной печени и не экспрессируется только терминально дифференцированными плазматическими клетками. Аналогично CD19 экспрессируется другой пан-В- маркер - CD72 , являющийся контррецептором CD5 , но он пока мало изучен.
Следующий этап дифференцировки - пре-В-лимфоциты - определяется, главным образом, по цитоплазматической экспрессии мю-цепи иммуноглобулина. На этом же этапе начинается экспрессия (слабая) CD20 и, по-видимому, CDw78 . CD20 - еще один пан-В-маркер, как и CD19 , часто использующийся для идентификации B-клеток. Параллельно появляется CD21 . Начало поверхностной экспрессии IgM свидетельствует о появлении незрелых B-клеток. Одновременно начинается поверхностная экспрессия CD22 , на предыдущих этапах обнаруживающегося только в цитоплазме. Примерно в это же время на поверхности B-клеток появляется еще несколько антигенов - CD37 , CD39 , CD40 . На поверхности незрелых B-клеток обнаруживается также ряд дифференцировочных антигенов: CD73 , CD74 , CDw75 и CD76 . Следующий этап - зрелые или покоящиеся B-клетки характеризуются одновременной экспрессией поверхностных IgM и IgD . Параллельно с IgD экспрессируется CD23 .
Дальнейшая дифференцировка проходит в периферических клетках крови или лимфоидных органах и вызывается антигеном. Она характеризуется увеличением размеров B-клеток и повышением уровня экспрессии антигенов MHC класса II . Это стадия активированных B-клеток. Антигензависимая дифференцировка вызывает замену поверхностных IgM/IgD другим изотипом (который будет позднее секретироваться) и деление, что свидетельствует о вступлении в стадиюB-бластов , или пролиферирующих B-клеток . Последние могут дифференцироваться либо в плазматические клетки, либо в B-клетки памяти . Плазматические клетки теряют поверхностную экспрессию большинства специфических B-клеточных маркеров (в том числе поверхностный Ig). Однако они опять начинают экспрессировать CD38 и, кроме того, сильно отличаются от B-клеток морфологически.
Процесс созревания и дифференцировки B-клеток , особенно последние его стадии, не всегда одинаково подразделяется на этапы.
В-л. Образующиеся в костном мозге иммунологически незрелые, поскольку они еще не поверглись воздействию АГ. Начальные этапы сохревания В-л. Не зависят от АГ. Пре-В-клетка временно продуцирует терминальную дезоксинуклеотидтрансферазу и общий АГ острого лейкоза(ОАОЛ;CD10). Несколько позднее экспрессирует характерные поверхностные АГ CD19, CD20[CD19 ( В4 ) - это гликопротеин, молекулярная масса которого равна 95 кДа. Полипептидная цепь состоит из 540 аминокислот. СD19 - экспрессируется на В-клетках ; мол. масса 95 кД; выполняет функцию корецептора.СТРУКТУРНАЯ ХАРАКТЕРИСТИКА. Внеклеточная область состоит из двух Ig-подобных доменов, разделенных областью, содержащей два остатка Cys. Эта область не имеет гомологии в аминокислотной последовательности с какими-либо известными белками. Большой цитоплазматический участок консервативен у разных видов млекопитающих и содержит несколько потенциальных мест фосфорилирования и пять потенциальных мест N- гликозилирования.ФУНКЦИИ. CD19 экспрессирован на всех В-лимфоцитах человека и на предшественниках В-клеток, но не на плазматических клетках. CD19 встречается также на фолликулярных дендритных клетках . CD19 включен в регуляцию В-клеточной пролиферации . Перекрестное связывание CD19 молекул без участия Ig ингибирует повышение концентрации свободных ионовкальция в цитоплазме и пролиферацию, индуцируемую антииммуноглобулиновыми антителам. CD20 ( В1 , Рр35 ) - это фосфопротеин, молекулярная масса которого равна 33 - 37 кДа. Полипептидная цепь состоит из 297 аминокислот. CD20 экспрессируется на В-клетках ; возможно участвует в активации В-клеток. СТРУКТУРНАЯ ХАРАКТЕРИСТИКА. Молекула содержит четыре трансмембранных сегмента. С и N-концы молекулы расположены внутри клетки. Фосфорлирование CD20 увеличивается в активированных клетках. CD20 обнаруживает гомологию с бета-цепью Fc-эпсилон-R1 . Общая организация структуры CD20 подобна структуре каналобразующих белков . ФУНКЦИИ. CD20 экспрессирован у человека и мышей только на В- лимфоцитах . У человека он встречается как на покоящихся, так и на активированных В-лимфоцитах, но отсутствует на плазматических клетках. CD20 принимает участие в В-клеточной активации и В-клеточной пролиферации . Ряд моноклональных антител к CD20 ингибирует клеточную пролиферацию, вызванную анти-Ig. В клетках Jurkat , трансфецированных геном CD20, этот белок непосредственно регулирует вход кальция в цитоплазму. Предполагают, что он формируеткальциевый канал .] и образует интрацитоплазматические μ-цепи иммуноглобулина. Когда В-л. Созревают, они экспрессируют на своей поверхности целые молеулы АТ. Последующие этапы созревания В-л. Зависят от АГ. С помощью Т-хелперов и специализированных макрофагов, антигенперзентующих, В-кл пролиферируют и созревают. Образующиеся в результате этих процессов плазматические клетки продуцируют большое количество иммуноглобулиновых молекул строго определенной специфичности. Характерный внешний вид: эксцентричное ядро с распредеоенным по периферии хроматином, базофильная цитоплазма, светлая чистая перинуклеарная зона с активным комплексом Гольджи. Другие стимулированные В-л. Становятся клетками долговременной памяти, сохраняющими информацию о ранее встречавшемся АГ, они быстро пролиферируют и продуцируют юольшое количество иммуноглобулина при повторной ыстрече с известным АГ.
Существуют 5 основных классов иммуноглобулинов IgG, IgA, IgM, IgD, IgE. Наиболее распространены IgG, есть 1,2,3 и 4. IgA имеет 2 подтипа: сывороточная и сереторная – находится в секретах слизистых и подслизистых, Ig D и IgE- минорные группы иммуноглобулинов, учавствующие в аллергических реакциях и реакциях гиперчувствительности замедленного типа. IgM полимеризуется, формируя большие пентамерные структуры.

Активация B-клеток вызывается либо неспецифическими поликлональными активаторами, либо перекрестным связыванием иммуноглобулиновых рецепторов одновременно с получением сигнала от макрофага или T-хелпера , распознающего номинальный антиген в комплексе с молекулами MHC класса II . Таким образом, B-лимфоциты реагируют на три различных типа антигенов:
Антигены тимус-независимые типа 1Некоторые антигены, такие, как бактериальный липополисахарид , при достаточно высокой концентрации способны к поликлональной активации значительной части популяции B-лимфоцитов , т.е. для такой активации антигенная специфичность поверхностных рецепторов клетки роли не играет.При низкой концентрации подобных антигенов, не приводящей к поликлональной активации, те B-лимфоциты, у которых иммуноглобулиновые рецепторы специфичны по отношению к данным антигенам, будут пассивно фокусировать их на своей поверхности. При этом за счет собственной митогенной активности эти антигены будут стимулировать пролиферацию клеток.Таким образом, тимус-независимые антигены типа 1 стимулируют деление B-клеток, взаимодействуя не с иммуноглобулиновыми рецепторами, а с другими структурами поверхностной мембраны.Тимус-независимые антигены вызывают преимущественный синтез IgM , и индуцируемый ими иммунный ответ практически не сопровождается формированием клеток памяти.
Антигенны тимуснезависимые 2 типа.Некоторые линейные антигены, медленно распадающиеся в организме и имеющие часто повторяющуюся, определенным образом организованную детерминанту , например, полисахарид пневмококков или полимеры D- аминокислот способны непосредственно без участия T-клеток стимулировать B-лимфоциты , т.е. относятся к тимус-независимым антигенам. Они длительное время персистируют на поверхности специализированных макрофагов краевого синусалимфатического узла и маргинальной зоны селезенки . Связывание этих антигенов с антигенспецифическими B-клетками происходит с высокой авидностью и обусловлено как перекрестным взаимодействием антигенных детерминант с иммуноглобулиновыми рецепторами (рис.6.13б ), так и вспомогательными факторами, выделяемымыми макрофагами . Таким образом, тимус-независимые антигены типа 2, по-видимому, вызывают деление клеток как за счет перекрестного связывания иммуноглобулиновых рецепторов, так и с помощью вспомогательных факторов, выделяемых макрофагами. Тимус-независимые антигены вызывают преимущественный синтез IgM , и индуцируемый ими иммунный ответ практически не сопровождается формированием клеток памяти .
TD (антигены тимусзависимые)Антигены T-зависимые (или тимус-зависимые) - это антигены, не способные непосредственно, без участия T-клеток стимулировать B- лимфоциты . Большинство природных антигенов является тимусзависимыми . Это означает, что полноценное развитие специфического иммунного ответа к таким антигенам начинается только после подключения T-лимфоцитов . Эти антигены в отсутствие T-лимфоцитов лишены иммуногенности: они могут быть одновалентными в отношении специфичности каждой детерминанты , подвергаться быстрой деградации фагоцитирующими клетками, наконец, не обладать собственной митогенной активностью. Связавшись с B-клеточными рецепторами, они, так же как и гаптены , не способны активировать B- клетку. Гаптены приобретают иммуногенность при соединении с подходящим белком-носителем . В настоящее время известно, что функция носителя заключается в стимуляции T-хелперов , помогающих B-клеткам реагировать на гаптен, стимулируя последние дополнительными сигналами ( рис. 6.10 ). Подобные представления сложились на основании опытов как in vivo, так и in vitro.