|
|
НЕЙРОХИМИЯ ПОДКРЕПЛЕНИЯ
После всего, что уже сказано, остается вопрос, указывает ли описанное выше сходство процессов индукции и подкрепления на сходство их механизмов' С моей точки зрения, да. Какие же факты это подтверждают? Какие нервные процессы начинают действовать во время индукции?
Для этого требуются анатомические пути, функционирующие во многих областях мозга, чтобы запустить в действие подкрепляющий «индуктор», химические агенты, стимулирующие реци-пиентную ткань к дифференциации. Пример подобного любопытного синтеза мы находим в нейрохимическом исследовании, проведенном Л. Стайном (рис. XV-1, XV-2).
Рис XV-1 Схема медиального переднемозгового пучка (верхний рисунок — система запуска) и перивентрикулярной системы волокон (нижний рисунок — система остановки) в мозгу млекопитающих (сагиттальный разрез) (Stein, 1968, Le Gros Clark et al, 1938)
«Все приведенные факты отвечают представлению о том, что механизм подкрепления содержит систему адренэргических Синапсов, которые в высшей степени чувствительны к фармакологическим воздействиям Усиление порадренэргической передачи в этих сииаптических аппаратах вызывает эффекты облегчения в поведении, ухудшение норадренэргической передачи подавляет поведение При всех условиях эти синаптические аппараты являются важнейшим участком работы мозга, где амфетамин и хлорпромазин оказывают влияние на целенаправленное поведение
Где локализуются эти синапсы? Сели в медиальном переднемозговом п\чке действительно находится система вознаграждения, как эю уьазыва.-лось выше, то данные адренэргические синапсы, очевидно, уже описаны 1руппой сотрудников Каролинскою института (Fuxe, 1965, Hillarp, Vuxe and Dahlstrom, 1966) Используя гистохимическую технику для обнар>же ния катехоламинов на клеточном уровне, эти исследователи сообщают
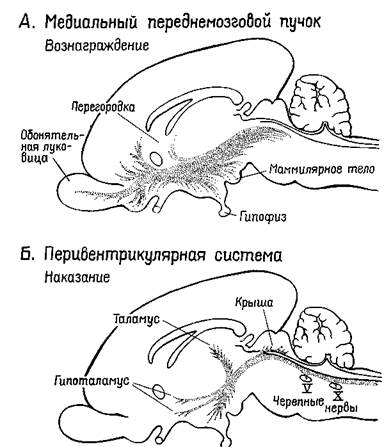

Рис. XV-2. Схема медиального переднемозгового пучка (пунктир) и пе-ривентрикулярной системы волокон в горизонтальной плоскости. На схеме гипоталамус представлен как три продольно расположенных столба — перивентрикулярный поясной слой, медиальная зона и латеральная зона. Пучки волокон в перивентрикулярной и латеральной зонах примыкают к медиальной зоне, которая лишена крупных пучков волокон, но содержит несколько известных ядер. Наклонные линии обозначают систему свода (Stein, 1968; Sutin, 1966).
о системе нейронов, содержащих порадреналин; клеточные тела этих нейронов берут начало в мезенцефальных отделах мозга, а волокна их восходит к медиальному иереднемозгопому пучку и оканчиваются в адренэргиче-ских синапсах гипоталамуса, лимбической доли и неокортекса... Используя совершенно иную технику, независимо от предыдущих авторов Геллер, Сайдеи и Мур (1966) подтвердили существование этой восходящей системы

Рис. XV-З. Ростральные проекционные пути неспецифической талампческой системы, идущие через нижние ножки таламуса (часть медиального переднемозгового пучка) в кору. Аксоны от передних неспецифических полей, включающих парацентральное ядро (PC) и, возможно, медиальную часть вентрального переднего ядра (VA), проходят рострально через нижние ножки таламуса как неспецифические проекционные пути (nsp) и через медиальный сектор каудального отдела полосатого тела (Str) к основанию орбитофронтальной коры (Cof). Эти аксоны широко ветвятся в субгризеальном белом веществе, и некоторые из них лродолжаются в ядрах ограды —п. accumbens (nAcSp). Остальные сокращения означают: ретикулярное ядро таламуса (nR), столбы свода (fxc), ограда (Sp) и радиация Цукеркэндла (rZ). Эпидермальная нейроглия (g) тянется вдоль боковых желудочков. Горизонтально-наклонное сечение, модифицированная методика быстрого окрашивания по Гольджи. 50-дневный, частично демиелинизированный мозг мыши. X150. В репродукции масштаб сокращен до Х0.75 (Scheibel, 1967).
волокон. Эти исследователи разрушали медиальный переднемозговой пучок па одной стороне на уровне латерального гипоталамуса и затем проверяли изменение содержания норадреналина в различных областях мозга. Поскольку в медиальном переднемозговом пучке аксоны в большинстве случаев не перекрещиваются, оказалось возможным сравнить уровень содержания норадреналина на пораженной и интактной сторонах. Было обнаружено, что на пораженной стороне — и только в структурах пучка мозга — норадреналина было значительно меньше; контрольные разрушения медиального гипоталамуса не привели к существенному уменьшению норадреналина. Эти результаты подтверждают существование в медиальном переднемозговом пучке волокон, содержащих норадреналин; кроме того, авторы приходят к выводу, что эти волокна входят в восходящую систему, так как уменьшение уровня норадреналина наблюдается только в структурах, расположенных выше уровня поражения...» (Stein, 1968, р. 110.)
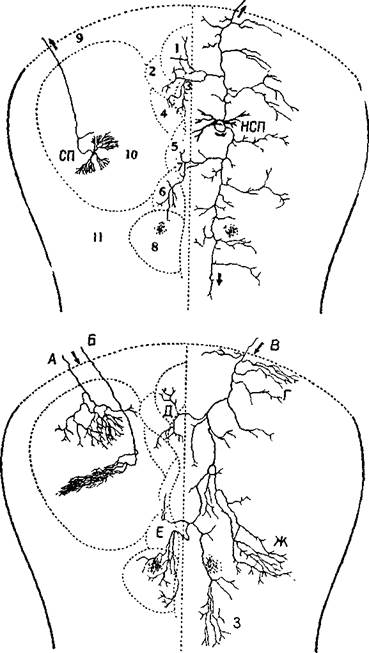
Шейбел и Шейбел (1967, рис. XV-3 и XV-4) также описали эту систему нейронов; по мнению этих авторов, она функционирует как система неспецифических афферентных путей к коре больших полушарий. Характерно, что аксоны этих клеток длинные и на их окончаниях имеются разветвления, обвивающие апикальные дендриты пирамидных клеток, свободно соединяющиеся с дендритами, в противоположность хорошо ограниченным синапсам, типичным для специфических афферентов. Эти данные, а также эксперименты Фукса, Гамбургера и Хокфельта (1968), показавшие, как отмечает Стайн, что неспецифические соединения являются в большинстве случаев адренэргетическими (то есть, что аксоны и их клетки содержат норадреналин), говорят
Рис. XV-4. Сравнение организации аксонных элементов специфической и неспецифической таламических систем. Верхний рисунок — таламофугальные элементы. Нейрон специфического вентробазального комплекса ядер (СП) посылает вертикально направленный без коллатералей аксон (10) к коре, исключение составляет лишь одна маленькая возвратная ветвь. Нейрон неспецифической системы (НСП) посылает аксон, который разветвляется на ростральные и каудальные отростки, богато снабженные коллатералями и идущие ипсилатерально и контр-латерально. Нижний рисунок — нисходящие таламопетальные элементы. А и Б — аксоны, идущие от корковых нейронов к комплексу различных специфических вентральных ядер. А — характерная объемная пограничная структура; Б — плоский древовидный нейрон, имеющий форму диска; В — аксон, идущий из коры к неспецифическим структурам. В диффузную, богатую коллатералями систему входят ветви, идущие к области ретикулярного ядра и к вентральному переднему ядру (Г), к неспецифическим структурам другого полушария (Д и Е), к заднему комплексу ядер (Ж) и мезодиенцефальным образованиям (3). Цифры обозначают: 1 — parataential; 2—anterior ventral; 3 — interanteromedial; 4 —r anterior medial; 5 — paracentral; 6 — central lateral; 7 — central medial; 8 —> centre median-parafascicularis complex; 9 — n. reticularis. Рисунок составлен на основании изучения ряда срезов мозга крыс и мышей до методу Гольджи (Scheibel, Scheibel, 1967).
о том, что эта система нейронов ответственна за запуск индуктивных процессов посредством стимуляции РНК и облегчения синтеза протеина. Стимулирующее влияние норадреналина на синтез, протеина, по-видимому, опосредствовано циклом адениномонофосфата (см. «The Role of Cyclic AMP in the Nervous System» in «Neurosciences Research Program Bulletin», July 1970, vol. 8). Кити как-то отметил: «Интересно, что стимуляция протеи-нокиназы посредством адениномонофосфата может значительно усиливаться с помощью ионов магнезии или калия и тормозиться кальцием, что показывает, каким образом эффект адренэргической стимуляции может по-разному воздействовать на умеренно активные и неактивные синапсы!»
Я подчеркнул бы еще и особое значение нейронной матрицы, на которой реализуется аминэргический подкрепляющий эффект. Эта матрица (нейронная сеть), чувствительная главным образом к холинэргическим веществам, и создает то, что в гл. XIV описывалось как компетентность мозговой ткани. Существуют данные, говорящие (рис. XV-5) о том, что адренэргическая стимуляция оказывает влияние на холинэргический механизм или по крайней мере — на активирующие и тормозящие механизмы мозгового ствола. Вспомним также, что в экспериментах Креча, Розенц-вейга и Беннета (рис. П-6) было обнаружено, что ацетилхолин участвует в эффекте влияния обогащенной среды на рост коры. Могут ли запускаться эти более продолжительные холинэргиче-ские процессы с помощью адренэргического механизма подкрепления или эти процессы протекают независимо от последнего?
Гликман и Шифф (1967) только на основе обширного обзора результатов, наблюдаемых в экспериментах по изучению влияния электрической стимуляции мозга на поведение, дали частичный ответ на этот вопрос. В этом обзоре выделяется два типа врожденных ответов, один из них состоит в «приближении», а другой в «удалении» от объектов-стимулов. Авторы установили корреляцию между участком электрической стимуляции в стволе мозга, откуда запускается подобный тип поведения, и положительным и отрицательным подкрепляющими эффектами при электрическом самораздражении (см. рис. V-1 и V-2). Они также высказали предположение относительно некоторых механизмов, посредством которых контролирующие кортикальные влияния (тормозящие и облегчающие) могут регулировать работу центрального эффекторного механизма ствола мозга. Однако они не поняли важности проблемы модификации поведения посредством подобных контролирующих корковых влияний. Это объясняется тем, что авторы не располагали фактами, которые указывали бы на вариабельную природу механизмов, приводящих к реакции (гл. XIV), в связи с чем возникает необходимость в особом, способном к модификациям состоянии, имеющем интегративные свойства. В настоящее время анатомическая основа подобных спо-

Рис. XV-5. Схема, показывающая возможные взаимосвязи между механизмами вознаграждения и наказания, составленная на основе экспериментов Маргулиса, Стайна и др. (1967, 1968) по химической стимуляции. Стимулы вознаграждения «запускают» поведение, освобождая его от тормозящих механизмов с помощью перивентрикулярной системы (ПВС), причем имеет место следующая последовательность явлений: 1) Активация медиального пе-реднемозгового пучка (МПП) с помощью стимулов, предварительно ассоциированных с наградой (или избежанием наказания) приводит к высвобождению норадреналина в амигдале и других подавляющих зонах переднего мозга (ПЗМ). 2) Тормозное действие норадреналина подавляет активность, сокращая тем самым опосредованное холинэргином возбуждение медиального таламуса и гипоталамуса (МТ и МГ). 3) Ослабление холинэргической синапти-ческой передачи в МТ и МГ уменьшает активность перивентику-лярной системы и таким образом снижает ее тормозное влияние на моторные ядра ствола мозга (Stein, 1968).
собных к модификации интегративных процессов найдена благодаря открытию системы адренэргических афферентов, берущих свое начало в стволе мозга и достигающих апимальных дендритов. коры больших полушарий.
Факты, на которых основываются гипотезы о сущности процесса индукции, получены почти исключительно в опытах на животных. Однако недавний обзор проблемы мозговых поражений у человека, вызывающих серьезные нарушения процессов памяти, неожиданно подтвердил существование механизма, подобного тому, который здесь описан. Ограниченные поражения коры больших полушарий (включая гиппокамп) или базальных ганглиев не всегда коррелируют с нарушениями памяти. Однаког когда патологический процесс затрагивает структуры ствола мозга, у больных наблюдается «неспособность к образованию.

Рис. XV-6. Сравнение транскортикальной негативной вариации (TNV) — в лобной, прецентральной и постцентральной областях коры обезьяны-ре-зус во время выполнения различных заданий на ожидание. В ситуации предъявления двух сигнальных стимулов, требующих двух ответов, обезьяна должна нажимать на рычаг в пределах 400 мсек после включения светового стимула, держать рычаг опущенным в течение двух серунд и отпускать его в пределах 400 мсек после включения звука. В ситуации предъявления одного сигнального стимула и двух ответов обезьяна должна была спонтанно начинать опускание рычага без стимула, держать рычаг опущенным в течение двух секунд и быстро отпускать его при включении звука. В ситуации отсроченных реакций свет и звук предъявлялись с интервалом в 2 сек; обезьяна должна была воздерживаться от ответа до тех пор, пока не появится звук. Каждая запись — усредненные компьютером 40—50 ответов. В качестве подкрепления обезьяны получали за каждый правильный ответ маленькие кусочки пищи. Ср. с рис. XV-7 (Donchin et al.,
1971).
новых следов памяти, то есть антероградная амнезия (когда продолжительное заучивание не помогает)», несмотря на «сохранность активного состояния внимания, уровня бодрствования-мозга» и «нормальную способность мыслить, решать задачи и т. д.»; в таких случаях при выздоровлении интеллектуальная деятельность лишь «немного ухудшается по сравнению с нремор-бидным уровнем, несмотря на грубые нарушения памяти». Анатомически «ядра, разрушенные поражением, во всех случаях располагаются по средней линии, а именно в 2—3 мм от эпендимы. Иными словами, их расположение в зоне третьего желудочка (и сильвиева водопровода) оказывается более важным как фактор, определяющий их чувствительность, чем все другие свойства этих ядер» (Adams, 1969, р. 98—102).
Нейрофизиологи также внесли известный вклад в изучение мозговых механизмов, определяющих процессы индукции как у человека, так и у животных. Несколько лет назад Грей Уолтер (Walter et al., 1964; Walter, 1967) проанализировал медленно протекающие изменения электрической активности мозга в различных ситуациях. Он обнаружил отрицательное отклонение электрического потенциала при действии одного сигнала, подготавливающего организм к Ответу на следующий, ожидаемый стимул. В наших экспериментах (Donchin et al., 1971; рис. XV-6 и XV-7) подобные отрицательные отклонения вызывались с помощью разнообразных подготовительных состояний ожидания и место возникновения изменений потенциала зависело от типа задания, требующего этого подготовительного состояния, например: если обезьяна или человек готовится, ничего не делая, к~ выполнению ответа, максимальная CNV (контингентная негативная вариация) появляется в лобной коре; если требуется подготовиться к моторному ответу, отрицательный потенциал появляется прежде всего в моторной коре и предшествует осуществлению движения; если требуется выполнить длительное ответное действие, например продолжительный подготовительный нажим на рычаг, максимальный отрицательный потенциал возникает в со-матосенсорной коре. Когда же наконец появляется ожидаемый второй стимул, отрицательный потенциал завершается резким положительным отклонением. В других экспериментах в ситуациях выработки оперантных условных рефлексов эти завершающие позитивные электрические потенциалы были связаны с появлением подкрепляющего агента (Clemente et al., 1964; Grandstaff, 1969; Lindsley, 1969). Таким образом, CNV с ее завершающим быстрым положительным потенциалом сигнализирует об ожидании-и его завершении, о подготовительных фоновых процессах, определяющих судьбу стимула, которые обсуждались в гл. XIV.
Непосредственный эффект может быть получен также с помощью отрицательной (катодной) и положительной (анодной) электрической стимуляции коры больших полушарий во время*
ЗИ

Рис. XV-7. Сравнение контингентных негативных отклонений (CNV) в различных участках мозга у человека в 5 различных экспериментах на время реакции. Во всех опытах предъявлялись 2 зрительных стимула с интервалом в 1,5 сек. В экспериментах А—В испытуемый должен был нажимать на кнопку большим пальцем ведущей руки в ответ иа второй стимул. г
В экспериментах Г и Д от испытуемого требовалось нажимать на кнопку сразу же после включения первого стимула, держать ее нажатой в течение интервала между стимулами и отпускать при появлении второго^ стимула. В качестве обратной связи использовался красный или зеленый свет, который зажигался после каждого правильного или ошибочного ответа. В экспериментах В — Д испытуемые должны были реагировать в пределах 350 мсек. В этих условиях ограниченного времени время реакции сокращалось настолько, что испытуемые допускали до 25% ошибок. Область отрицательных колебаний оценивалась методом суммирования всех отрицательных отклонений (относительно исходной средней линии), возникающих в течение двухсекундного интервала, следующего за включением первого стимула. Электронегативные мозговые волны оценивались, усреднялись и интегрировались с помощью компьютера системы LINC-8.
научения (Stamm, 1961; Morrell, 1961). Отрицательная стимуляция повышает скорость научения, положительная оказывает про-
•тивоположное влияние.
Все эти эксперименты дают убедительное доказательство того, что изменения электрического потенциала возникают в мозгу и
.до и во время подкрепления. Однако мы фактически ничего не
знаем о тех механизмах, которые вызывают эти изменения электрического потенциала. Столь же мало знаем мы и о нейрохимических процессах, сопровождающих эти электрические изменения. Возможно, что CNV отражает или мобилизует «холинэргиче-скую компетентность», а положительные потенциалы возникаюг вследствие активации адренэргических путей или вызывает эту активацию. Тот факт, что электрические изменения связаны с подкреплением, открывает пути для изучения данной проблемы на поведенческом, гистологическом и химическом уровнях — пути, которые в течение ряда лет были полностью закрыты. Понимание подкрепления как процесса индукции выдвигает гипотезы, которые могут быть проверены в биологических исследованиях и помогут разгадать ту загадку, над которой полвека безуспешно бились бихевиористы.