|
|
Анастасия З., поступила курс на первый курс в НТЦ ПНИ в 10 месяцев.
Развитие коммуникативных функций и их нарушения при аутизме и органических поражениях головного мозга
И.А.Скворцов, В.М.Башина
Коммуникация в обществе всегда подразумевает активное взаимодействие. Развитие коммуникативных функций ребенка проходит несколько этапов, формируются полимодальные анализаторные пути, как компоненты реализации коммуникативной деятельности. Коммуникативная деятельность всегда содержит как общение со средовым окружением, так и эгоцентрические процессы, уменьшающие внешнюю зависимость «Я» и ограждающие его от внешних воздействий. Поэтому и развитие анализаторов различной модальности включает импрессивные и экспрессивные компоненты: анализ экзогенной и эндогенной информации, а также ответную реакцию – внешнюю и внутреннюю, эгоцентрическую.
Развитие анализаторов в онтогенезе.Функции зрительного, слухового, тактильного и др. анализаторов, прежде всего, импрессивные. Однако без экспрессивной компоненты, которая наиболее ярко выражена в речевой функции, эти анализаторы не могли бы функционировать. Очевидно, существует специфический внутренний информационный «язык» анализатора. Так, аналогично «внутренней речи», существуют «внутреннее зрение», «внутренний музыкальный слух» и т.п. По аналогии с импрессивной речью существуют импрессивные зрение и слух – способность воспринимать зрительные и слуховые информацию и образы, а по аналогии с экспрессивной речью – экспрессивное зрение художника и экспрессивный музыкальный слух композитора.
Развитие нервно-психических функций ребенка контролируется гибкой генетической программой, предусматривающей возможные средовые воздействия на каждом онтогенетическом этапе и включающей соответствующие ответные реакции. Импринтинг, состыковка афферентно-независимых командных нейронов со средой через посредничество афферентно-зависимых и промежуточных нейрональных групп является важнейшим инструментом развития навыков специфического поведения (рис. 1).
Рис. 1. Взаимодействие командных и зеркальных нейронных систем в онтогенезе импрессивного и экспрессивного компонентов анализаторов.

Поведенческий шаблон, регулируемый командными нейронами, конкретного организма адаптируется к конкретной среде. Весь терминальный исполнительный аппарат (двигательный, перцептивный и др.) уже функционирует к моменту рождения для обеспечения реализации генетически детерминированных жизненно-важных поведенческих шаблонов, и интенсивно развивается в дальнейшем онтогенезе. Ребенок учится не сокращать отдельные мышцы, а выполнять и совершенствовать под влиянием среды «готовые» автоматизмы – генетически детерминированные комплексы двигательных реакций, использующие различные комбинации активности множества мышц и нервов.
Развитие поведенческих реакций (двигательных, перцептивных, речевых и др.) осуществляется с помощью обучающих систем мозга – зеркальных систем нейронов, обеспечивающих фиксацию, сопереживание и копирование поведенческих реакций окружающих, а также посредством деятельности мозжечковой и стриато-паллидарной систем, способствующих автоматации каждого этапа обучения всех анализаторных систем конкретного организма. Вначале автоматизмы носят общий, базисный характер, но постепенно под воздействием разнообразных воздействий среды и индивидуального опыта вырабатываются все новые и утонченные их варианты. автоматизмы становятся более конкретными, способными обеспечивать специфическую реакцию на любые «случаи жизни» (для любых условий имеются свои отдельные автоматизированные ответы). При этом программы реализации конкретных композиций автоматизмов размещаются в формирующихся корковых центрах гнозиса и праксиса.
В дальнейшем все большая автоматизация и конкретизация автоматизмов и их комплексов в повседневной деятельности человека (на уровне зрелого, а затем и инволюционного этапов онтогенеза) приводит к относительной функциональной незанятости и дистрофии (от бездействия) обучающих систем мозга, что усугубляет и делает необратимым инволюционный процесс. Творческий, созидательный потенциал обучающих систем мозга при этом угасает, активность их нивелируется, в результате они становятся более подверженными дегенеративным дистрофическим процессам, в том числе генетически детерминированным. Старение этих систем – один из компонентов общей и всеобъемлющей программы умирания организма, завершающей онтогенез.
Импрессивные и экспрессивные функции анализаторов.Формирование и работа основных анализаторов подробно изучены на примере зрительного восприятия. Созревание анализаторных функций и особенно рецепторно-периферического образного восприятия в значительной степени генетически детерминировано. Отсрочка, смещение начала развития зрительного и слухового восприятия на второй месяц жизни не случайны. Это позволяет приоритетно включить сразу после рождения жизненно-важные двигательные реакции сосания и шага.
Нарушения развития перцептивных функций, зрительного, слухового восприятия, как правило, сопровождают внутриутробную и перинатальную патологию мозга и наблюдаются в сочетании с нарушениями двигательного, интеллектуального, коммуникативного и речевого развития, реже в изолированном виде. Речь идет не только о врожденных слепоте и тугоухости, но, главным образом, о нарушении восприятия в целом, как сложного комплекса функционирования анализатора. На фоне реабилитационной терапии компенсаторные возможности могут открываться на самых разных уровнях анализатора: от рецепторно-периферического до коркового.
Вживление в область ганглиозных зрительных клеток сетчатки кролика множества микроэлектродов, позволяет регистрировать ответы не только на примитивные световые раздражения, но и на сложные (например, лицо человека) объекты (Верблин Ф., Роска Б., 2007; FriedS.I., MunchT.A., WerblinF.S., 2005; RoskaB., MolnarA., WerblinF.S., 2006). Последующая обработка полученных данных на компьютере позволяет получить «изображение» изменяющейся мимики лица последовательно каждую секунду. Отмечено, что практически «готовое» изображение лица формируется уже на уровне сетчатки, а не исключительно в проекционной и гностической зрительной коре, как предполагалось ранее. В проекционной коре суммированная информация от обоих глаз перекодируется, интегрируется и систематизируется, а затем поступает в гностическую область зрительной коры, где анализируется и сопоставляется с хранимыми ранее принятыми образами, которые при этом уточняются и совершенствуются, а сам гностический процесс все более автоматизируется.
Работа коркового отдела зрительного анализатора, как и других анализаторов, афферентно-независимо подготавливается командными системами нейронов внутриутробно и в течение первого постнатального месяца. Эта стадия зрительного онтогенеза генетически детерминирована и практически не зависит от средовых воздействий. Лишь в конце первого, в начале второго месяца жизни (третья-четвертая недели) к работе зрительного анализатора подключаются группы афферентно-зависимых и переходных нейронов, и появляется возможность их интеграции с группой командных нейронов.
Начинается новый этап – период интеграции со средой и формирования в коре отраженных зрительных образов. На этом этапе в развитии каждого анализатора и любой формы восприятия огромную роль играют зеркальные системы нейронов, которые будут рассмотрены ниже. Они обеспечивают «зеркальное» отражение, сопереживание воспринимаемой информации, а также феномен имитации – подражания. Способность к имитации – важнейший механизм развития психоневрологических функций, который включается, начиная с первых часов жизни, и имеет важнейшее значение в процессе копирования и обучения.
По-видимому, работа корковых блоков зрительного, слухового, тактильного, соматосенсорного и др. анализаторов имеет общую схему деятельности, в отличие от соответствующих рецепторных систем, которые существенно отличаются в принципе и жестко специфичны. Поэтому на фоне реабилитации образование специфического коркового анализатора возможно в неспецифическом месте, поскольку специфичность восприятия связана, главным образом, с рецепторно-периферическим уровнем. Кора относительно «голографична», то есть может нести в себе различную по модальности анализаторную специализацию, однако, эта «голографичность» обусловлена не только универсальностью корковых нейронов, но и универсальностью способа гностической обработки информации от различных специфических рецепторов.
Память и хранение специфических образов – динамичный процесс, поддерживаемый непрерывным функционированием кольцевых (афферентно-эфферентных) систем самоактивации образного восприятия и памяти. Кроме того, эфферентное обеспечение процесса гнозиса осуществляется обслуживающими двигательными системами (например, глазодвигательными системами, позволяющими фиксировать внимание на объекте, разглядывать и изучать его; системами слухового внимания, прислушивания, определения локализации источника звука и т.п.).
Особое значение имеют системы экспрессии образа (рис. 2), обеспечивающие произвольное внутреннее воспроизведение образа – вспоминание, непроизвольные эгоцентрические проекции образа (галлюцинаторные?), а также творческое воспроизведение образа, его материализация в среде в виде рисунка, скульптуры, музыки, поэзии, двигательных и позных «рисунков» в балете и др. Большинство из нас не претендует на лавры живописца или композитора, и врачи не лечат отсутствие у нас таланта художника или музыканта. А экспрессивной речью (тоже определенный талант от природы) должен овладеть каждый ребенок (хотя именно особым индивидуальным, генетически детерминированным талантом импрессивной и экспрессивной речи определяется непостижимая загадка поэзии Пушкина, прозы Гоголя).
Таким образом, анализаторная экспрессия (зрительная, слуховая, обонятельная, вкусовая, тактильная, речевая и др.) включает: произвольное воспроизведение (вспоминание) образов во внутреннем зрении, внутреннем слухе, внутренней речи и т.п. (знакомого лица, знакомого голоса, вкуса и запаха пищи, конкретного прикосновения и др.); воспроизведение образов в рисунке, мелодии, движении (например, при копировании мимики и жестов, увиденных у другого человека, имитации голоса и др.); воспроизведение различных эгоцентрических действий.
Рис. 2. Импрессивный и экспрессивной компоненты функционирования анализаторов.

Функционирование анализатора обеспечивается специфическим рецепторно-периферичес-ким аппаратом, а также корковыми импрессивными и экспрессивными системами, тесно связанными между собой на всех уровнях интеграции. Работа как импрессивных, так и экспрессивных компонентов анализатора на ранних этапах генетически детерминирована, а затем в ходе онтогенеза становится все более зависимой от среды и адресованной среде. Кроме того, мозговые структуры, обеспечивающие гнозис, находятся в тесном взаимодействии с соответствующими центрами праксиса. Однако гностические центры, афферентные, по сути, имеют и собственное эфферентное обеспечение, так же как и праксические центры, эфферентные, по сути, имеют собственное афферентное обеспечение.
Генетическая программа, через командные системы нейронов подготавливает автоматизированный некоммуникативный стереотип импрессивного и экспрессивного действия, который трансформируется в коммуникативное, адресованное среде поведение лишь по мере укрепления афферентных связей со средой, развития анализаторных систем, функций восприятия. Дальнейшее развитие анализаторов, активное обучение в общении со средой происходит при участии зеркальных нейронных систем.
Роль зеркальных нейронов в коммуникации.В начале 90-ых годов прошлого века итальянский исследователь Джакомо Риззолатти описал так называемые командные «зеркальные системы нейронов», с которыми связана способность к «сопереживанию» и повторению и, следовательно, к обучению под влиянием окружения.
У здоровых людей ПЭТ и МРТ регистрируют активизацию этих нейрональных групп как при выполнении определенных действий, так и при наблюдении за такими действиями других людей. «Сопереживание», внутреннее повторение увиденного или услышанного без движения – основа способности к подражанию, в том числе эмоциональному, поведенческому, речевому и т.п. зеркальные системы нейронов, расположены в различных отделах коры мозга, но, главным образом, – в прецентральной (в области речевого центра Брока), нижне-фронтальной, поясной, островковой, нижне-теменной областях, угловой извилине (рис. 3). Они обеспечивают «зеркальное» отражение, сопереживание воспринимаемой информации, а также феномен имитации – подражания. Дисфункции зеркальных нейронов островковой и передней поясной коры могут обусловливать неспособность к сопереживанию, а нарушения зеркальной системы угловой извилины – дефекты речи.
Имеется ЭЭГ-показатель повышения активности зеркальных нейрональных групп. Импульсная активность зеркальных нейронов премоторной коры подавляет мю-волны в электроэнцефалограмме (частота мю-ритма составляет 8-13 Гц.). В норме, когда человек совершает некое движение, регистрируются импульсы, которые генерируют моторные нейроны. В отличие от них зеркальные нейроны премоторной коры генерируют импульсные разряды и при выполнении движений, и в том случае, когда человек наблюдает за выполнением действия другим лицом. Одновременно подавляются мю-волны.
Исследователи регистрировали мю-волны у детей с аутизмом и контрольных испытуемых, когда они совершали произвольные движения или наблюдали за такими же действиями по телевизору (во время просмотра видеоклипов с изображением сжимающейся и разжимающейся кисти). Дети сжимали и разжимали правую кисть руки, что вызывает подавление мю-волн в ЭЭГ как при аутизме, так и контрольных испытуемых. Однако при наблюдении за аналогичными действиями других людей мю-волны подавлялись только у здоровых детей (амплитуда мю-волн у контрольных испытуемых резко уменьшалась, а у детей с аутизмом подавление мю-ритма отсутствовало). Полученные результаты свидетельствуют о нарушении работы зеркальной системы мозга у детей с аутизмом преимущественно в нижней фронтальной извилине – одном из отделов премоторной коры мозга. Данным обстоятельством можно объяснить их неумение распознавать намерения других людей. Способность к имитации – важнейшая особенность нервной системы, которая прослеживается, начиная с первых часов жизни, и имеет важнейшее значение в процессе обучения. Зеркальные системы нейронов относительно неспецифичны и участвуют в развитии всех форм анализаторной деятельности: зрительной, слуховой, соматосенсорной, речевой и др.
Рис. 3. Функциональные области коры левого полушария и зоны зеркальных нейронных систем в мозге (по В.Рамачандран, Л.Оберман, 2007).

Полагают, что в основе таких признаков аутизма, как отсутствие сопереживания, игнорирование и отвержение средовых воздействий, лежит дефект или снижение активности зеркальных нейронных систем. При аутизме нарушение или аберрация чувства сопереживания, а также неспособность распознавать цели и намерения действий и переживаний других людей связаны с отсутствием активизации зеркальных нейронных систем при наблюдении за действиями и намерениями других. у больных с аутизмом она наступает только при непосредственном выполнении действий. Отсутствие «чувства сопереживания», возможно, определяет появление и других специфических признаков аутизма, таких как стереотипные эгоцентрические поведенческие и речевые реакции, замыкающиеся «на себя» при отсутствии регистрации и сопереживания окружающих событий.
Очевидно, что у больных с аутизмом эгоцентрические эквиваленты универсальны и должны иметь место в работе всех анализаторов – зрительного, слухового и др., но, в отличие от речевой функции, они могут быть менее яркими. Чрезвычайно интересны сообщения о высоком творческом потенциале у лиц, наблюдавшихся в детстве в связи с аутизмом и различными эгоцентрическими проявлениями, равно как и о «странностях» и отчужденности у известных ученых, художников, артистов.
Рис. 4. Экспрессивные функции сенсорных двигательной и речевой систем. Роль зеркальных нейронных систем и реакций имитации.

В онтогенезе развитие функциональной активности зеркальных нейронов тормозит эгоцентрическую, генетически детерминированную автоматизированную деятельность, которая постепенно угасает. Так, по мере развития подражательной речевой активности формируется коммуникативная экспрессивная речь, а эгоцентрическая автоматическая речь подавляется. По-видимому, в переходе от автоматизированной деятельности, обусловленной командными нейрональными системами, к связанным со средой поведенческим реакциям определенную пусковую и кондукторную роль играют зеркальные нейроны и развитие механизмов подражания (рис. 4). В дальнейшем онтогенезе механизмы подражания постепенно тормозятся развитием произвольной деятельности, но не угасают совсем. В неблагоприятных условиях возможна диссолюция – растормаживание реакций подражания и одновременно – эгоцентрических проявлений.
Существенным отличием человеческого общения от взаимодействия животных – это возможность не только знаковой, но и речевой коммуникации. При этом зеркальные нейроны, достигающие именно у человека высшей степени организации играют важную роль в развитии коммуникации ребенка (Rizzolatti, G. et al., 1996; Rizzolatti G., Craighero L., 2004; Rizzolatti G., Fadiga L., Fogassi L., Gallese V., 2002). В речевом онтогенезе до определенного момента у ребенка функционируют эндогенные механизмы, организующие вокализации, но в период появления осознанных слов должно включаться произвольное подражание звуковым комплексам извне – имитация. Е.Н.Соколов (1981, 2003) связывал развитие речи с включением и развитием систем зеркальных нейронов, благодаря которым осуществляется функция имитации, копирования и обучения. Активация этих нейронных структур, позволяет воспроизводить поведение, аналогичное наблюдаемому.
Речевой анализатор в коммуникации, импрессивная и экспрессивная речь. Способность к имитации особенно необходима в ходе развития экспрессивной речи (не случайно зеркальные нейроны концентрируются в области центра Брока). Лишь при нормативном развитии как импрессивной, так и экспрессивной речи формируется коммуникативная речевая функция. Экспрессивные функции не менее сложны, чем импрессивные, но определенным образом зависят от последних, как бы базируясь на них, и окончательно созревают позднее, хотя эгоцентрическая экспрессивная речь может проявиться до завершения формирования импрессивной речи.
Процесс формирования речи имеет генетически детерминированную компоненту, и на ранних стадиях афферентно-независим от среды и некоммуникативен (эгоцентричен), хотя экспрессивная речь направлена, прежде всего, на коммуникацию между людьми. Эта начальная стадия развития экспрессивной речи является эгоцентрической – в значительной степени автоматической и в определенном смысле – внутренней. Эгоцентрическая речь нередко опережает в своем появлении развитие полноценной импрессивной речи. Экспрессивная эгоцентрическая речь неэффективна в общении, поскольку не является коммуникативной и, по-видимому, определяется патологической персистенцией афферентно-независимой автоматической речи, связанной с командными системами нейронов, своевременно не получившими средового подкрепления. Эгоцентрическая речь напоминает «автоматическое» бесполезное сосание у дезафферентированных новорожденных котят. В условиях патологии, при детском аутизме эгоцентрическая речь не редуцируется, доминирует в речевой экспрессии и сопровождается игнорированием-отвержением речевого контакта с окружающими.
Дальнейшее становление экспрессивной речи под влиянием определенного речевого окружения или, напротив, блокирование ее развития связывают с работой зеркальных систем нейронов, обеспечивающих способность подражания чужой речи и обучения языку. Способность к имитации особенно необходима в ходе развития экспрессивной речи (не случайно зеркальные нейроны концентрируются в области центра Брока). Экспрессивная речь не менее сложна, чем импрессивная, но определенным образом зависит от последней, как бы базируясь на ней, и окончательно созревает позднее (хотя эгоцентрическая экспрессивная речь может проявиться уже на ранних стадиях развития импрессивной речи). Лишь при нормативном развитии как импрессивной, так и экспрессивной речи формируется коммуникативная речевая функция.
Коммуникативные функции: восприятие и сознание, активная и пассивная коммуникация.Связь с окружающим миром, окружающей средой присуща всему живому, включая не только животных, но также и растения. В самом примитивном виде она включает внешнее дыхание, регулирование температуры, обмен веществ. Сам по себе рост, увеличение объема животного, растения, человеческого тела – своеобразное внедрение, захват территории среды. Взаимосвязь между особями животного мира характеризуется появлением стайного, стадного сообществ. Как стая, так и стадо не просто группировки большего или меньшего множества животных. В той или иной степени они действуют как единое целое, подчиняясь общим «побуждениям». Стая – не сумма животных, она становится отдельным организмом. Для каждого животного стая является отчасти внешней средой, но для целой стаи отдельная особь – ее неотъемлемая часть, при этом понятие внешней среды значительно расширяется (внешняя среда по отношению к сообществу).
Любое взаимодействие животных особей подразумевает некоторое ущемление или ограничение индивидуальных действий, но предоставляет новые возможности совместного взаимодействия со средой. Взаимодействие в стае отчасти нивелирует индивидуальное значение отдельной особи, которая перестает быть полностью самостоятельной. Поэтому такое сообщество подразумевает взаимодействие двух противоположных побуждений: стремление особи к автономности, независимости от среды и, с другой стороны, стремление в стаю, стремление быть частью, членом стаи. Сообщество животных напоминает безмасштабную сеть с элементами-концентраторами. Коммуникация – внутрисетевая функция. в процессе включения и закрепления в сети новый элемент может пройти путь от рядового сетевого элемента до элемента-концентратора.
В человеческом обществе коммуникативные взаимодействия значительно сложнее и претерпевают существенные преобразования по мере развития ребенка, значительно более медленного, чем у животных. Для ребенка взаимодействие в человеческой среде подразумевает и более выраженное стремление к интеграции в обществе и более активное воздействие среды на него. В связи с этим представляет интерес модель «Маугли» – развитие ребенка среди животных. Нарушение развития человеческой речи у человека в животной среде – результат суррогатного для человека «общества» – нечеловеческого окружения. Однако животная среда не только не способствует развитию человеческой речи у «Маугли», но активно блокирует формирование речи (не свойственной животным) у человеческого ребенка, которого она приняла («интегрировала») в себя. Воздействие окружающих людей «втягивает» ребенка в интеграцию, одновременно обучая его коммуникативным функциям. Коммуникация – не только индивидуальная функция ребенка, но и популяционная функция общественной среды.
Следует рассматривать пассивную и активную, импрессивную и экспрессивную коммуникацию. Импрессивная коммуникация базируется на генетически детерминированной коммуникативной потребности, восприятии и осознании предлагаемого контакта, «восприятии интеграции» своего «Я» в человеческом обществе. экспрессивная коммуникация (проекция генетической программы на среду) – это осознание себя в человеческом обществе (желание, ожидание и мотивация контакта с окружающими людьми), активный поиск, развитие взаимодействия в обществе, совершенствование автоматизмов установления межличностного контакта, стремление к лидерству.
Коммуникативные действия, деятельность, операции, направленные на взаимодействие с обществом в целом или отдельным человеком могут быть и не внешними, а только внутренними (аналогично внутренней речи). Коммуникация без внешних действий – внутреннее ощущение и осознание контакта, себя, своего «Я» в контакте. В норме коммуникация способствует формированию и утверждению своего личностного образа, как члена общества, самовыражению «Я» в коммуникации. Поэтому коммуникативный образ человека является своеобразным его портретом (или автопортретом).
Как отмечает А.Г.Рузская (1999), «ребенок вступает в общение с окружающими людьми… без потребности в общении… Коммуникативная потребность не врожденна. Она возникает в первые два месяца жизни на основе первичных органических нужд детей и их потребности в новых впечатлениях». С этим трудно согласиться, поскольку именно генетически детерминированное и врожденное «знание» о среде (еще никогда не встречавшейся во внутри- и внеутробной жизни), ее «ожидание» и, главное, адресованность среде специфических поведенческих автоматизмов, сформированных афферентно-независимыми командными системами, лежат в основе универсального феномена импринтинга. Адекватность человеческого окружения, открывшегося пришедшему в мир ребенку, его генетически детерминированным «ожиданиям» является важным фактором дальнейшего развития коммуникативной деятельности.
Описанный нами ранее (Скворцов И.А., 2001) феномен экспансии функциональных систем на среду лежит в основе формирования коммуникации. Среда и, в особенности, человеческая среда не является чисто внешним фактором, постоянное взаимодействие с ней индивида делает ее своеобразной дополнительной функциональной системой организма, коммуникативным соучастником. Развитие пассивной и активной коммуникации, с одной стороны, генетически детерминировано (это – коммуникативная потребность, самоощущение в среде и распространение своего «Я» на среду), а, с другой стороны, – результат активного воздействия на ребенка человеческой среды и «втягивания» его как личности в коммуникативную сеть человеческого общества (рис. 5).
взаимодействие ребенка с окружающей человеческой средой всегда связано с компромиссом: коммуникативная потребность ребенка требует его приспособления к законам, правилам, условностям, образу действия, принятым средовым обществом, и, соответственно, сопровождается некоторым ущемлением личностных потребностей. Однако среда не просто принимает, а активно вовлекает ребенка в себя, одновременно обучая его общению и заставляя выполнять средовые условности и требования. Важно отметить взаимное стремление: у ребенка – захватить в себя среду, а у среды – захватить в себя ребенка. Среда поглощает личность, но интеграция в среде дает возможность индивиду воспринимать окружающее и реагировать на него не с индивидуальной позиции, а с коллективной? воспринимать себя как часть Целого общества (семейного, национального, общечеловеческого).
Важнейшее значение в развитии у детей навыков общения имеет взрослый, который постоянно «подтягивает» деятельность ребенка на новый, более высокий уровень развития. Обогащение содержания коммуникативной деятельности детей и их взаимоотношений с окружающими приводит к смене устаревших форм общения новыми, что стимулирует психический прогресс ребенка. Организуемая взрослым практика взаимодействия с детьми способствует преобразованию их социальных потребностей. Без постоянной поддержки взрослого развитие общения замедляется или даже прекращается. Активное вмешательство взрослого вызывает благоприятные сдвиги в общении детей, устраняя дефекты и отклонения.
Рис. 5. Условная схема биологических основ коммуникации, взаимодействия ребенка и среды.

Внешним признаком инициации коммуникативных контактов считается взгляд в глаза с характерной мимикой ожидания ответного взгляда. Содержательность коммуникативных контактов, их количество, интенсивность зависят от возраста детей, уровня психического развития. В общении со взрослым у детей последовательно возникают возрастающие коммуникативные потребности: в 2–6 мес. ребенку требуется доброжелательное внимание; в 0,5–3 года – сотрудничество; в 3–5 лет – уважительное отношение взрослого; в 5–7 лет – взаимопонимание и сопереживание.
Уже в конце первого года жизни появляются специфические для отношений со сверстниками действия: повышенная эмоциональность и особая непринужденность, свобода действий. На 3-м году жизни возрастает потребность в общении со сверстниками. В общении со сверстниками ребенок требует: в 2–4 года – соучастия партнера в забавах, его внимания и доброжелательности; в 4–6 лет – сотрудничества и признания ровесником, сопереживания и взаимопонимания. Коммуникация оказывает влияние на психическое состояние сверстников. Радостное чувство, возникшее при первых взаимодействиях с другими детьми, в дальнейшем будет окрашивать в радостные тона все, с чем встретится ребенок, а конфликт приведет к возникновению негативизма, подавленности, агрессии.
Наряду с развитием коммуникативной деятельности постепенно формируется образ «Я». первичным сенситивным периодом для появления положительного эмоционального самоощущения является первое полугодие жизни ребенка, период позитивного эмоционального общения. Возникает первичное отражение ребенком самого себя, которое складывается из представления о себе и связанными с ним переживаниями. Образ «Я» не является постоянным на протяжении жизни. Он начинает складываться в раннем онтогенезе под влиянием индивидуального опыта взаимодействия, общения с окружающими людьми и постоянно изменяется и развивается в онтогенезе. Под влиянием индивидуального опыта формируется позитивная оценка окружения и положительное самоощущение, принятие, осознание интеграции себя в общественной среде.
Сознание, как коммуникативная функция.Сознание является существенным фактором становления коммуникативных функций. В физиологической природе сознания, как важнейшей функции мозга, остается еще много неясного. S.A.Greenfield (2000, 2005), также как C.Koch (2004) рассматривают сознание в тесной связи с восприятием. Восприятие предусматривает осознание воспринимаемой информации по нескольким модальностям – зрительное, слуховое, тактильное осознание и т.п. Однако сознание – это не только восприятие и не просто осознание восприятия отдельной модальности, это восприятие присутствия себя в мире, которое относительно независимо от наличия или отсутствия какой-либо из перцептивных модальностей. Оно может быть сохранено у слепых, глухих, слепоглухонемых, а также, в определенном виде, – у больных аутизмом при отвержении коммуникации по основным анализаторным каналам. Развитие сознания невозможно без восприятия, но сформированное сознание уже не столь зависит от отдельного вида восприятия. Вместе с тем, для самого восприятия изначально необходимо наличие сознания, готовности к восприятию.
Сознание контролирует взаимодействие мозга как с внешней, так и с внутренней средой, в том числе и во время сна. Сон сопровождается снижением уровня сознания, но не полным выключением его (о чем свидетельствует память о сновидениях и осознание их при пробуждении). Это снижение дает возможность мозгу отдохнуть от напряженного состояния бодрствования. Оно же позволяет проявляться сновидениям. Мозг переходит на иной режим работы (как бы на «автопилот»), в основном связанном с внутренними церебральными и висцеро-церебральными циркуляциями сигналов и межсистемными вертикальными и горизонтальными циркуляциями в самом мозге. Возможно, что подобные сновидениям процессы протекают и в бодрствующем мозге, но не осознаются. Последовательное угнетение сознания при углублении наркоза или постепенном входе в коматозное состояние также сопровождается более или менее яркими сноподобными ощущениями различной, чаще смешанной модальности (чувство умиротворения, яркий свет, тепло, другие слуховые, тактильные и висцеральные ощущения).
Сознание – это функция целого мозга, а не отдельных анализаторов, оно обеспечивается при участии стволово-подкорково-корковых кольцевых систем, нервных ансамблей, ответственных за уровень активности мозга, функции сна и бодрствования, за включение пробуждения и засыпания. Сознание – это не состояние, а процесс, непрерывное функционирование. Нервные ансамбли «создают сознание» только когда они действуют, функционируют, то есть меняются, «живут». Замедление или остановка этого процесса нарушает или выключает сознание. Нарушения сознания могут быть обусловлены полным или частичным блокированием обеспечивающих его систем, при их повреждении, а также вызываться различными функциональными изменениями – расстройствами сна, психической регуляции уровня бодрствования, адекватности перцепции и внимания.
Таким образом, функция сознания имеет как качественные, так и количественные характеристики. Вместе с тем, трудно провести четкую грань между нулевым и минимальным сознанием. Может ли быть нулевой уровень сознания у живого человека? Каковы реальные, а не «договорные» границы состояния смерти мозга?
В клинике неврологии развития нередко трудно различать коммуникативные нарушения и различные формы расстройств сознания. В особенности это касается раннего детского возраста, когда коммуникация и сознание еще формируются. При тяжелых морфологических дефектах мозга у детей раннего возраста (МРТ-признаки гипоплазии или дисплазии коры, мозолистого тела, гипоплазии мозжечка и ствола мозга и др.) врожденная недостаточность развития паллиума (как аналог апаллического синдрома взрослых) может имитировать различные задержки или аномалии становления коммуникации, в частности, при раннем детском аутизме, аутистических синдромах.
В условиях текущего структурного и функционального созревания мозга чрезвычайно сложно разграничить сами понятия «сознание» и «коммуникация», поскольку коммуникация возможна только при наличии сознания, а само сознание, по сути, является отражением коммуникативной потребности, включая и осознание своего «Я» в коммуникативном взаимодействии со средой. Наличие сознания необходимо для формирования коммуникации, при его нарушениях коммуникация невозможна или становится неполноценной. Аналогично этому коммуникация нарушается и при расстройствах перцепции. В раннем возрасте при структурном дефекте срединных и вертикальных стволово-подкорково-корковых систем, а также при нарушении развития восприятия коммуникация формируется с отклонениями или аномально. различные формы детского аутизма составляют особую группу коммуникативных нарушений, патогенез которых связывают, в частности, с дефектом зеркальных нейронов мозга.
Как видно на представленной схеме (рис. 6), нарушения коммуникации могут быть обусловлены как поражениями структур ЦНС, так и задержками развития коммуникативных функций. К первой группе относятся нарушения сознания и сна, психо-эмоциональные расстройства и органические поражения головного мозга. Эти нарушения не носят врожденного характера и наблюдаются преимущественно у взрослых и детей старшего возраста. Задержки коммуникативного развития возникают при врожденных гипоплазиях структур головного мозга, при первичных нарушениях перцептивного развития или общей недостаточности формирования психоневрологических функций и нередко имитируют нарушения коммуникации при поражениях мозговых структур. Врожденная аномалия коммуникативного развития характерна для раннего детского аутизма и аутистических синдромов.
В раннем детском возрасте особенно очевиден переход от пассивной коммуникации (способности позитивно воспринимать коммуникативные воздействия окружающих) к активной (активный полимодальный поиск коммуникативного контакта). В дальнейшем происходит непрерывное совершенствование навыков общения (рис. 7, 8). Развитие как пассивной, так и активной коммуникации, совершенствование их протекает при непосредственном участии человеческого окружения (побуждение к коммуникации, обучение общению). Взаимодействие эндогенной потребности в коммуникации и активирующего влияния человеческого окружения является важнейшим фактором развития коммуникативных функций у ребенка.
Рис. 6. Условная схема коммуникативных расстройств у детей при нарушениях развития психоневрологических функций.
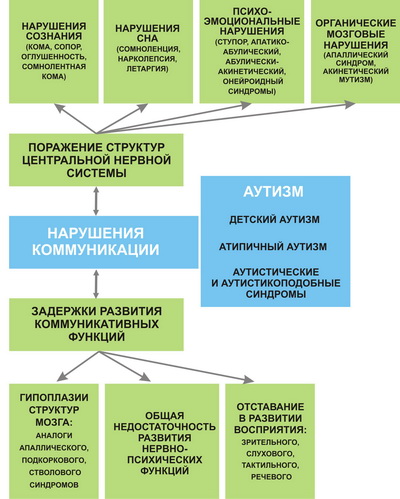
Рис. 7. условная схема формирования пассивной и активной коммуникации.
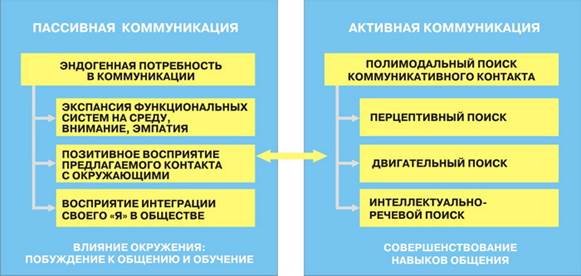
В основе развития пассивной коммуникации лежат: врожденная генетически детерминированная потребность в коммуникации, феномен экспансии функциональных систем на среду, внимание, сопереживание и эмпатия, позитивное восприятие контакта с окружающими. Сопереживание – переживание состояния, подобного эмоциональному состоянию другого человека может быть непроизвольным, неосознаваемым или осознаваемым. Эмпатия – внерациональное познание человеком внутреннего мира других людей (вчувствование), а также и осознанная эмоциональная отзывчивость на переживания другого (по механизму децентрации). Первичные эмпатические реакции наблюдаются уже у новорожденных, в дальнейшем они усложняются, обобщаются, когнитивизируются и становятся переживаниями, определяющими отношение ребенка к другим людям.
Вместе с тем, большую, нередко определяющую роль играют мотивационные установки, врожденное позитивное или негативное отношение к внешним контактам. При позитивной установке ребенок стремится к интеграции себя в общественной среде, постепенно получая все большее удовлетворение от общения, от ощущения себя частью общества. Напротив, при негативной установке общение, развитие коммуникативных функций блокируется: или временно (при задержках развития того или иного восприятия – реакции игнорирования-отвержения по определенной модальности), или составляют стержень дальнейшего развития ребенка (при детском аутизме и аутистических синдромах).
Рис. 8. Эндогенные и средовые факторы развития пассивной и активной коммуникации.

На первой неделе после рождения отсутствие реакций на окружающее является не только проявлением послеродового стресса в ответ на сверхпороговую силу зрительных, слуховых, тактильных и висцеральных раздражений, но и своеобразной негативной «коммуникативной» установкой с неприятием среды, в которую вступил новорожденный. В дальнейшем в первую очередь нивелируется негативные тактильные реакции. На втором месяце, по мере созревания и интеграции структур зрительного и слухового анализаторов ребенок переходит от характерного для первого месяца жизни негативного неприятия внешних звуков и света к постепенному развитию адекватного перцептивного восприятия. Начало активной коммуникации в норме во втором триместре приходит на смену пассивной коммуникации, и период игнорирования внешней среды практически полностью завершается. Считается, что для детей первого полугодия жизни характерно ситуативно-личностное общение – генетически первая форма общения ребенка со взрослым.
При аутизме, основным признаком которого лежит нарушение коммуникации, коммуникация не отсутствует вовсе, а извращается, является эгоцентрической, односторонней, изолированной. Однако игнорирование и отвержение подразумевают все же наличие регистрации окружающего мира и, нередко, дифференциацию человеческого и неодушевленного окружения.
Общие сведения об аутистических расстройствах(В.М.Башина). Эволюционно-биологическое, клинико-психопатологическое, динамическое, катамнестическое исследования и лечение когорты детей (более 7000 человек) с разными видами аутистических расстройств легли в основу анализа состояния проблемы аутизма (НЦПЗ РАМН, 1984–2008 гг.). В последнее десятилетие произошел отказ от феноменологического определения «ранний детский аутизм, шизофренического спектра» Каннера. В отдельных странах стали использоваться другие его верификации: аутистический синдром (Rutter М., 1978; АРА, 1987; детский аутизм (WingL., 1980); болезнь Каннера (Ковалев В.В,1995). Согласно МКБ – 10 пересмотра ВОЗ (1994, 1999) и Классификации аутизма НЦПЗ РАМН признано выделение ряда аутистических расстройств: детский аутизм, детский процессуальный аутизм, атипичный аутизм, атипичный детский психоз, УМО (умственная отсталость) с аутистическими чертами, синдром Аспергера, синдром Ретта (РС) и др.
Аутистическое расстройство (АР) – есть психическое расстройство, с проявлениями дизонтогенеза, что отражено и в его двойном названии: «Первазивное (общее) нарушение психологического развития. Аутистическое расстройство». АР, как сложное клиническое образование, нельзя отнести ни только к психическим симптомам, ни исключительно к нарушениям развития. Базисные проявления этого вида патологии у детей – есть аутизм, как психическое (психотическое) расстройство и дизонтогенез, как нарушение развития.
Клиническая картина классического детского аутизма (ДА), эндогенного генеза представлена аутистическими психическими признаками – особыми аномалиями в психическом взаимодействии, в виде отвержения общения, речи, и симптомами сбивки, расщепления в функционировании всех систем организма, нарушениями в онтогенезе – в виде сочетания соответствующих возрасту форм в деятельности, со смешанными, дезинтегративными видами искаженного, задержанного, или вовсе остановившегося развития во всех функциональных сферах организма. Дезинтегративный тип дизонтогенеза, отнесен нами к следствиям расщепления в деятельности высших психических функций с переходом на подкорковые виды функционирования, которые свойственны преимущественно аутистическим симптомокомплексам, аутистическим расстройствам эндогенного генеза, шизофренического спектра. Функционирование на подкорковом уровне наиболее отчетливо в моторной сфере ребенка-аутиста: в атетозоподобных движениях в пальцах рук, в стереотипной, повторяющейся деятельности, общем поведении и монотонной эмоциональности.
При ДА на разных этапах его течения и возрастного онтогенеза (2–15 лет и далее), в 3/4 случаев выявлен периодический переход от позитивного эволютивного типа течения к негативному прогредиентному, процессуальному. Глубина процессуальности вариирует в степени выраженности: от вялого и более выраженного злокачественного непрерывного течения до приступообразного течения с подострыми и острыми психотическими эпизодами, приступами.
В возникновении атипичного аутизма (АА) при УМО генного, хромосомного, обменного, экзогенного генеза, подтверждена гетерогенная почва. Атипичный аутизм представлен разными видами течения: вялым непрерывным, вялым с обострениями, подострым, острым психотическим. Клиническая картина аутистических черт в круге АА с вялым и подострым течением, лишь частично соотносится с аутизмом при ДА. В ней присутствует отрешение, избегание, однако почти не определяется явлений психической сбивки в функционировании, нет психического «расщепления», свойственного эндогенному аутизму. В этих случаях можно говорить о фенотипическом сходстве отдельных черт аутизма. На отдаленных этапах АА с подобным вялым течением признаки аутизма нередко нивелируются, несмотря на продолжающееся нарастание первоначального базового процесса. Наблюдаемые явления фенотипического сходства отдельных признаков аутизма в структуре АА с гетерономной почвой нередки. Это явление по И.В.Давыдовскому (1964) отнесено – к эквифинальности. А.В.Снежневский (1970) рассматривал его, «как результат экономии приспособительных механизмов организма».
Другая группа АА с чертами аутизма, может протекать подостро или остро в форме психозов, близких по клинической картине к атипичным психозам (эндогенного круга) и процессуальному аутизму с психотическими проявлениями расщепления, нарушениями, сбивкой в функционировании. Такой тип АА с чертами аутизма, протекающего в форме психозов, полагаем, близок к эквифинальным «экзогенным реакциям» по Бонгофферу, к группе «периодических психозов» по А.В.Снежневскому (1970). Вопрос о природе этих атипичных психозов у детей, остается открытым. Ранее в психиатрии эти стертые аутистические признаки неэндогенного генеза, возникающие на гетерогенной почве, определяли «аутистикоподобными», в МКБ-10 пересмотра (1994) их отнесли к группе «атипичного аутизма». По поводу гетерогенности природы атипичного аутизма (генной, хромосомной, экзогенной) уместно привести слова А.В.Снежневского: «Существующие сейчас нозологические единицы не являются окончательными, требуется еще большая, кропотливая работа по изучению всех особенностей течения процессов и их закономерностей. В результате будет преодолен жесткий нозологизм, будет преодолено и принятие синдрома за нозологическую единицу».
В аспекте обнаруживаемых нейроморфологических нарушений аксонодендритных связей – при синдроме Ретта с аутистическим начальным периодом (Беляченко П.В., 1996) и другой патологии перивентрикулярных областей мозга, например, при УМО с аутистическими чертами на органической почве (Скворцов И.А., 2003) можно увидеть некое подтверждение возможности эквифинальности в круге атипичного аутизма (АА) при поражении неоднозначных структур мозга. Черты аутизма в клинике АА – его первичность, вторичность, коморбидность несомненно многообразны. В круге АА сам признак аутизма может возникать при нарушении развития в связи с основным расстройством – при синдроме Ретта, синдроме Дауна, туберозном склерозе, фенилкетонурии и др. При АА признак аутизма может появляться вследствие сочетанности базового расстройства и психоза (возможно эндогенного круга) и в таких случаях возникает сходство не в связи с эквифинальностью, а в связи с эндогенной структурой психотического процесса. Последние цитоморфологические исследования корковых структур, дают возможность высказать версию, что при эндогенном аутизме блокада информации связана с синаптическими расстройствами в коре головного мозга.
При органических поражениях головного мозга – по причине нарушения проводящих путей в перивентрикулярных областях – также страдает приток информации. При парааутизме экзогенного происхождения, когда нет блокирования информационного поля, а есть недостаточность информации за счет обеднения социальной среды, приток информации также уменьшается. Разные причины нарушения информационного поля во всех случаях качественно меняют состояние областей-мишеней, что в целом способствует формированию черт аутизма и задержке умственного развития ребенка.
При определении клинической структуры аутистических расстройств (АР), прежде всего, оценивается уровень развития ребенка в доманифестном периоде: нормальный онтогенез или дизонтогенез – стигмированного, искаженного, задержанного, дезинтегративного типов. Оценивается сущность АР, как психического аутистического нарушения в виде отторжения взаимодействия с окружающим миром, погружения в свой собственный мир. Констатируются проявления психического расщепления – дезинтегративные черты в функционировании и другие признаки нервно-психического дизонтогенеза. Во всех случаях АР корковая, продуктивная, развивающая деятельность ребенка частично замещается подкорковой примитивной, стереотипной. Характер этой деятельности взаимосвязан с глубиной наступающего нейродизгенеза. Новые данные о наличии в критические периоды онтогенеза (роста) ребенка явлений физиологического апоптоза, изменения клеточных популяций нейронов в центральной нервной системе, их особом взаимодействии с областями-мишенями, нуждающимися в наибольшей стимуляции моторной, речевой, эмоциональной и других функциональных систем, помогают понять механизмы формирования аутистических кризовых возрастных периодов в онтогенезе. Именно на эти периоды в большинстве случаев и приходится манифестация аутистических проявлений, АА.
При АА формирование черт аутизма может идти вяло на почве постепенной равномерной утраты корковых функций (в связи с базисным расстройством), без явлений дезинтегративности. Поэтому в этих случаях можно говорить лишь о фенотипическом сходстве аутистических черт, без расщепления функционирования. В этих случаях утрата функций опережает проявления аутистических черт, создает базу отвергания, как утрату функционирования, без черт расщепления. В случаях коморбидности АА (УМО при синдроме Дауна, туберозном склерозе и др.) с аутистическими психозами, которые протекают с явлениями расщепления, аутистические симптомы выражены более отчетливо.
Все рассмотренные аутистические расстройства (АР) в зависимости от этапа их становления, глубины расстройства, коморбидности с психозами – имеют свои отличительные клинико-психопатологические особенности, течение и исходы. Дифференциация АР особенно важна для подбора терапии и применения коррекционной педагогической реабилитации. Истинную причину аутистических расстройств с явлениями расщепления, дезинтегративного дизонтогенеза у детей, не удалось узнать за прошедшее столетие, несмотря на заметные подвижки в понимании их патогенеза. В большинстве случаев АР признается геннная патология. В патогенезе АР всегда присутствует нарушение целенаправленного поведения, падение активности, в группе ДА эндогенного генеза – сбивка в функционировании, нарушение психического «включения» в корковый высший круг деятельности.
Филогенетические аспекты коммуникативных функций в норме и при аутизме.в самой коммуникации (особенно на ранних стадиях онтогенеза человека) заложен компонент негативизма, отвержения, сопротивления вовлечению в полимодальные контакты; активная коммуникация обеспечивается функционально более высоким интегративным уровнем мозга, компромиссным осознанием необходимости общения, порождающим радость коммуникации. У животных механизмы коммуникации безусловны, и свойственный человеку конфликт между «общественным» и «личным» у них отсутствует. Есть ли аналоги негативизма у животных? Несомненно у них присутствуют эгоцентричность, отчужденность, отсутствие чувства юмора, которые являются своеобразными средствами ограждения от внешней среды, но аутистические негативизм и отвержение, по-видимому, свойственны только человеку-аутисту. Можно предположить, что состояния анабиоза у животных, связанные с определенными критическими атмосферными, погодными и др. явлениями, с отказом от функционирования, временным переходом на примитивное функционирование, «замирание» – близки к аутизму.
Особенности моторики, характерные для детского аутизма, отражают филогенетически и онтогенетически более ранние или «архаичные» способы поведенческого реагирования: в движениях рук – манипуляция с последними (ульнарными) пальцами кисти и игнорирование первых трех (медианных), осуществляющих захват щепоткой; в движениях ног – ходьба на носках – не из-за спастики икроножных (большеберцовых) мышц, а в связи незрелостью функциональной системы перонеальной группы мышц. первые три пальца кисти и перонеальные мышцы являются «чисто человеческими», обеспечивающими «собирание мелких предметов щепоткой» и тыльное сгибание стопы при вертикальной ходьбе. На более ранних стадиях филогенеза – у животных (даже у способных на вертикальную ходьбу) – эти двигательные системы функционально не развиты.
чем являются аутистические дисфилогенетические и дизонтогенетические признаки – проявлением феномена диссолюции, возврата назад или задержкой, застреванием на промежуточной стадии функционального формирования нервно-психических функций. Несомненно, задержкой, «застреванием». Задержка развития на этих архаичных для человека двигательных стереотипах характерна для детей с аутизмом и аутистическими синдромами. По мере формирования психоневрологических функций (пусть аномального) эти архаичные стереотипы «вкрапливаются» в процесс формирования функций гнозиса и праксиса, закрепляются в образной памяти двигательного и перцептивного поведения и становятся необратимыми, препятствуя дальнейшему нервно-психическому развитию.
На фоне активной реабилитационной терапии улучшение в состоянии детей с аутизмом и аутистическими синдромами протекает не по нормативному пути, ряд аутистических проявлений нивелируется или видоизменяется, но общие поведенческие особенности нередко длительное время сохраняют аутистические черты, что дает основание говорить о так называемом «леченном аутизме». Процесс «ухода» от аутистического эгоцентризма имеет особенности. В речи – при преобладании эгоцентрической речи коммуникация нередко начинается с шопота, наблюдается эгоцентрическая речь как бы безразлично, безадресно обращенная к собеседнику, а также «вкрапление» эгоцентрических эмболов во вновь создающуюся коммуникативную речь.
В поведенческих реакциях – первые активные двигательные, перцептивные коммуникации ребенок начинает производить через посредника, чаще через мать, манипулируя ее рукой для захвата, указания, прикосновения или отталкивания, наблюдается эгоцентрическое по внешним проявлениям, но целенаправленное по смыслу действие, то есть сохраняется замкнутость при элементах осознанного действия. Так, в семье с двумя детьми с атипичным аутизмом при синдроме Мартина-Белл старший мальчик, прошедший несколько курсов лечения в НТЦ ПНИ с заметным улучшением в клинике, заботливо следил и ухаживал за больной младшей сестрой при сохранении у него признаков эгоцентрического, отчужденного поведения. Соотношение сравнительной выраженности коммуникативных нарушений и показателей интеллектуального развития в клинике детского аутизма может быть очень вариабельным: удельный вес того и другого причем интеллектуальный уровень детей-аутистов может быть резко сниженным до уровня УМО средней или тяжелой степени тяжести (напр., при синдроме Каннера, синдромах Мартина-Белл, Ангельмана, Дауна и др.) или, напротив, весьма высоким, особенно в отдельных, специфических областях (известны аутисты среди видных ученых, музыкантов, артистов).
Настороженность, недоверие к среде характерно для начала развития восприятия любой модальности и проявляется в виде неспецифических неаутистических реакций игнорирования-отвержения (Скворцов И.А. с соавт., 2003). Эти реакции наблюдаются также при задержке или аномалии развития восприятия. При этом важна степень недостаточности прихода в корковые центры модальной информации, а причина недостаточности может быть связана с любой локализацией блока прохождения импульса по вертикальным путям анализатора. Например, для зрительного анализатора причиной реакций игнорирования-отвержения может быть недостаточность самого глаза и его сетчатки, зрительных нервов, четверохолмия и наружных коленчатых тел, зрительного пучка в височной доле, проекционного и гностического центров в затылочной доле.
Реакции возникают на грани перехода от полного отсутствия восприятия до минимального восприятия и носят протопатический характер. Протопатическое восприятие всегда имеет негативный отвергающий оттенок, но все же отражает минимальную способность ребенка реагировать на модальные воздействия. Ребенок не чувствует и не осознает зрительного, слухового или тактильного воздействия, но все же негативно реагирует на него, значит, какое-то минимальное восприятие присутствует. В зрительном и слуховом восприятии ребенок не видит и не слышит, но реагирует негативно, он как бы «видит, что он не видит» и «слышит, что он не слышит». Протопатическая реакция игнорирования-отвержения является, по-видимому, универсальной и на последовательных этапах онтогенеза лежит в основе различных феноменов.
Таким образом, нарушение коммуникативных функций у детей может быть обусловлено различными причинами, они могут доминировать в клинике в виде проявлений детского аутизма или атипичного аутизма, аутистических синдромов или сопровождать нарушения двигательного, перцептивного развития при ДЦП, врожденном снижении зрительного, слухового восприятия и т.п. Полное или частичное сужение контактов с окружающими и особенно со сверстниками при тяжелой инвалидизации детей также приводит к задержке развития или аномалии коммуникативных функций.
Характерно, что при всех формах детского аутизма и аутистических синдромов, по данным МРТ-исследований, обнаруживаются относительно однотипные морфологические изменения в головном мозге, преимущественно в виде гипоплазии червя и полушарий мозжечка, а также гипоплазии полушарий и дисплазии коры большого мозга. При этом нарушения координации движений, как правило, отсутствуют, преобладают коммуникативные и когнитивные отклонения.
В лечении коммуникативных нарушений у детей акцент обычно делают на методах педагогической, психологической и логопедической коррекции, которую настойчиво и последовательно проводят в течение многих месяцев и лет. Мы исходили из представления о коммуникативных нарушениях у детей и, в частности, о детском аутизме, как о дизнейроонтогенетической патологии (что базируется на данных МРТ головного мозга у этих детей с гипоплазией коры большого мозга, мозолистого тела, червя и полушарий мозжечка и др.). Поэтому в комплексную реабилитацию была включена активная стимулирующая фармакотерапия, позволяющая активизировать формообразующие и регенеративные процессы в мозге и ускорять его развитие.
Таблица 1. Сравнительная эффективность лечения в НТЦ ПНИ детского аутизма Каннера и синдрома Аспергера (по: В.М.Башина с соавт., 2003).
| Исследуемые функции | Клиническая форма заболевания | |||||
| Детский аутизм Каннера | Синдром Аспергера | |||||
| +% | +% | |||||
| Тонкая моторика | 1,1 | 1,8 | 1,1 | 2,5 | ||
| Экспрессивная речь | 1,0 | 1,8 | 1,5 | 3,5 | ||
| Импрессивная речь | 1,6 | 2,1 | 1,4 | 2,9 | ||
| Запас знаний | 1,9 | 2,2 | 2,1 | 3,5 | ||
| Когнитивные функции | 5,6 | 7,9 | 6,1 | 12,4 | ||
| Ориентировочные реакции | 1,7 | 3,1 | 2,9 | 4,1 | ||
| Общение | 0,4 | 1,0 | 0,8 | 1,4 | ||
| Эмоциональная сфера | 0,9 | 1,6 | 1,1 | 2,1 | ||
| Коммуникативные навыки | 3,0 | 5,7 | 4,7 | 7,6 |
1 – оценка состояния функции до лечения, 2 – оценка состояния функции после лечения, +% - относительное увеличение показателя после курса терапии (в %).
В исследованиях В.М.Башиной (1999), В.М.Башиной и соавт. (2003) показана возможность эффективного лечения коммуникативных и когнитивных нарушений при детском аутизме с применением препаратов гидролизатов мозга (церебролизина, церебролизата), в частности, в ходе комплексной реабилитационной терапии, проводимой в НТЦ ПНИ (таблица 1).
***
В НТЦ ПНИ реабилитационное лечение проводилось по ранее разработанной нами комплексной методике (Скворцов И.А., 1993, 2000, 2008). Она включает метамерную фармакотерапию, направленную на стимуляцию нейронального метаболизма, которая осуществляется инъекционно или безыгольно (с помощью точечного микроэлектрофореза и глубокого фармакомассажа). Используемые при метамерной фармакотерапии препараты гидролизатов мозга и другие стимуляторы оказывают многостороннее влияние на нервную клетку и функциональные системы, активизируют синтез ДНК и белка в нейронах и структурах мозга и оказывают ростстимулирующее действие. Одновременно проводится форсированная сенсорная стимуляция основных анализаторных систем, способствующая воссозданию или восстановлению в функциональных системах мозга «образов» двигательных, зрительных, слуховых, речевых и других поведенческих реакций. Эти два компонента комплексной терапии неразрывно связаны: метамерная фармакотерапия создает потенцию, готовность мозговых структур и функциональных систем к перестройке, но создание нормативной функции взамен аномальной может произойти только путем освоения этими системами новых поведенческих образов.
За 19 лет работы в НТЦ ПНИ комплексную нейрореабилитационную терапию прошел 451 ребенок с коммуникативными нарушениями различной природы, включая детский аутизм и аутистические синдромы, коммуникативные нарушения при ДЦП и других нарушениях развития психоневрологических функций (НРПНФ). Как правило, больные проходили повторно 2-3 курса терапии (всего проведено 1114 курсов). Общие данные по пролеченным больным представлены в таблице 2.
Как видно из приведенной таблицы, больные с детским аутизмом, включая синдромы Каннера и Аспергера составили 52,1%, аутистические и аутистикоподобные синдромы – 36,5%. В 30,1% случаев коммуникативные нарушения разной степени тяжести сопровождали клинику ДЦП и других нарушений развития психоневрологических функций (НРПНФ), включая апаллическиподобный синдром (2,2%) при тяжелых структурных дефектах полушарий мозга.
В 1997 г. нами описано 44 ребенка с сочетанием ДЦП и аутизма (Скворцов И.А., Башина В.М., Ройтман Г.В., 1997) и 34 больных аутизмом без ДЦП, которые прошли лечение в НТЦ ПНИ. Среди больных с сочетанием клиники ДЦП и аутизма преобладали дети с атонически-астатической и атактической формами ДЦП (54,5%), спастические тетра- и гемиформы составили 34,1%, гиперкинетические и спастико-гиперкинетические – 11,4%.
Таблица 2. Больные с нарушениями коммуникативных функций, проходившие лечение в НТЦ ПНИ.
| Синдром | Больные | Курсы |
| Детский аутизм, ранний детский аутизм | 201 (44,6%) | 445 (39,9%) |
| Атипичный аутизм | 44 (9,8%) | 95 (8,5%) |
| Аутистический синдром | 108 (23,9%) | 181 (16,2%) |
| Аутистикоподобный синдром | 57 (12,6%) | 113 (10,1%) |
| Синдромы Каннера и Аспергера | 34 (7,5%) | 71 (6,4%) |
| Коммуникативные нарушения у детей с ДЦП и другими НРПНФ | 126 (27,9%) | 268 (24,1%) |
| нарушения коммуникации при аномалиях развития и грубых структурных дефектах мозга – апаллически-подобный синдром | 10 (2,2%) | 21 (1,9%) |
| Всего коммуникативных нарушений | 451 (100%) | 1114 (100%) |
Требует дальнейшего изучения вопрос, развивается ли синдром детского аутизма при детском церебральном параличе параллельно основному заболеванию как проявление сочетанного нарушения онтогенетического развития двигательных и коммуникативных функций или возникает вторично – в патогенетической связи с возникшими при ДЦП двигательными и психическими нарушениями, приводящими ребенка к инвалидности и изоляции. При рассмотрении сочетания ДА и ДЦП важно исходить из достоверной доказанности диагноза как ДЦП, так и аутизма, поскольку при детском церебральном параличе у больных нередко наблюдаются те или иные отклонения в коммуникативном развитии, а при разных видах ДА, как правило, отмечается задержка не только в психическом, но и в двигательном развитии, а также явления мышечной дистонии, которые могут ошибочно рассматриваться как начальные признаки ДЦП (например, как атонически-астатическая его форма).
Нарушения коммуникации по типу апаллически-подобного синдрома. У детей первых лет жизни при тяжелых врожденных дефектах полушарий головного мозга, кистозном перерождении мозгового вещества, перивентрикулярной атрофии или выраженной гипоплазии коры возможности коммуникации, пассивного и активного взаимодействия с окружающим миром существенно снижаются. Это особенно заметно в первые месяцы жизни, на фоне задержанного формирования основных психоневрологических функций. Ребенок практически не реагирует на тактильные, зрительные, слуховые в