|
|
Внутреннее строение
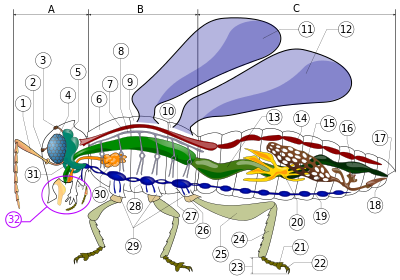
Морфология насекомого
A — голова, B — грудь, C — брюшко
| 1. антенна, 2. глазки (нижний), 3. глазки (верхний), 4. сложный глаз, 5. мозг (cerebral ganglia), 6. переднегрудь, 7. дорсальная артерия, 8. трахеи, 9. среднегрудь, 10. заднегрудь, 11. переднее крыло, 12. заднее крыло, | 13. средняя кишка (желудок), 14. сердце, 15. яичник, 16. задняя кишка (кишечник, прямая кишка и анальное отверстие), 17. анус, 18. вагина, 19. абдоминальныйганглий, 20. мальпигиевы сосуды, 21. подушечка, | 22. коготки, 23. лапка, 24. голень, 25. бедро, 26. вертлуг, 27. передняя кишка (пищевод, желудок), 28. грудной ганглий, 29. тазик, 30. слюнная железа, 31. подглоточный ганглий, 32. ротовой аппарат. |
Дыхательная система
Дыхательная система большинства насекомых представлена множеством трахей, пронизывающих всё тело и открывающихся наружу с помощью дыхалец (стигм), которые регулируют поступление воздуха. Трахейная система насекомых может быть открытой (типична для многих наземных свободно живущих видов или для водных насекомых, дышащих атмосферным кислородом) либо замкнутой. При втором типе трахеи имеются, но дыхальца отсутствуют. Этот тип характерен для водных или эндопаразитических насекомых, дышащих кислородом, растворённым в воде или в полостной жидкости хозяина. Многие водные личинки насекомых, имея замкнутую трахейную систему, дышат растворённым в воде кислородом и обладают жабрами, которые обычно располагаются по бокам брюшных сегментов или на кончике брюшка.
В открытую трахейную систему воздух поступает через дыхальца, количество которых меняется от одной — двух пар до восьми — десяти пар. Количество и места расположения дыхалец отражают приспособление насекомых к условиям мест обитания. Каждое дыхальце ведёт в атриальную полость, стенки которой образуют замыкательный аппарат и систему фильтрации воздуха. Трахеи ветвятся и опутывают все внутренние органы. Концевые ветви трахей заканчиваются звёздчатой трахейной клеткой, от которой отходят самые мелкие разветвления, имеющие диаметр 1—2 мкм (трахеолы). Их кончики лежат на клеточных оболочках либо проникают внутрь клеток. У многих хорошо летающих насекомых имеются воздушные мешки, представляющие собой расширения продольных трахейных стволов. Их полость не является постоянной и может спадаться при выходе воздуха. Воздушные мешки принимают участие в вентиляции крыловой мускулатуры и выполняют аэростатическую функцию, уменьшая удельный вес летающих насекомых.
Ткани внутренней среды
Жировое тело и гемолимфу рассматривают как единую систему тканей внутренней среды насекомых.

Личинка комара-звонца, в крови которой имеется дыхательный пигментгемоглобин
Гемолимфа является единственной тканевой жидкостью в теле насекомых. Она состоит из жидкого межклеточного вещества (плазмы) и форменных элементов (клеток). Плазма либо бесцветная, либо окрашена в желтоватый или зеленоватый цвет. Кровь личинок комаров-звонцов красноватого цвета из-за наличия гемоглобина, переносящего кислород пигмента. Содержание воды в гемолимфе 75—90 %. Плазма содержит неорганические соли, питательные вещества, ферменты, гормоны, пигменты. Реакция гемолимфы слабокислая или нейтральная (pH=6—7). Солёность гемолимфы соответствует 0,7—1,3 % раствору хлорида натрия.
Главная функция гемолимфы — снабжение органов питательными веществами и удаление из них продуктов обмена. Гемолимфа также содержит гормоны, выделяемые железами внутренней секреции и участвующие в регуляции физиологических процессов. Дыхательная функция гемолимфы незначительна и ограничена растворённым в ней кислородом. Гемолимфа также создаёт внутреннее давление в организме насекомого, поддерживая форму тела у мягкотелых (например личинок). Давление гемолимфы используется для осуществления некоторых видов механического перемещения, например, для расправления крыльев, развёртывания хоботка, разрыва покровов при линьке. У некоторых насекомых (саранчовые, кузнечики, нарывники, божьи коровки) гемолимфа выбрызгивается в целях самозащиты (автогеморрагия) или содержит биологически активные вещества и выделяется наружу при опасности. Важна функция иммунитета, которую выполняет гемолимфа. Иммунная система насекомых включает яды и антибиотики плазмы (например, кантаридин и насутин), белковые ферменты плазмы (лизоцим), а также амёбоидные гемоциты, которые осуществляют фагоцитоз одноклеточных паразитов и образуют гемоцитарную капсулу вокруг многоклеточных. У насекомых нет специфического иммунитета с образованием антител по типу иммунитета позвоночных. Однако у некоторых насекомых наблюдались случаи кратковременного специфического иммунитета, где роль антител играли вещества небелковой природы. При повреждении покровов у многих насекомых гемолимфа, вытекающая наружу, способна свёртываться и образовывать кровяной сгусток.
Жировое тело представляет собой рыхлое клеточное образование, не имеющее строгой локализации и расположенное внутри тела. Его функция заключается в накоплении запасов питательных веществ и участии в промежуточном метаболизме. Во время линьки, голодания, метаморфоза, при развитии половых продуктов идиапаузе происходит расходование резервных веществ, накопленных в трофоцитах жирового тела. Многочисленные насекомые, не питающиеся на стадии имаго, существуют за счёт запасов, накопленных в жировом теле на личиночных стадиях. Кроме трофоцитов, в состав жирового тела входят уратные клетки, накапливающие мочевую кислоту, и мицетоциты — специальные клетки, в которых живут симбиотические микроорганизмы.
У ряда насекомых имеется бактериом — специализированный орган, в котором находятся эндосимбиотические бактерии. Клетки органа обеспечивают питательные вещества и место обитания бактериям, в обмен те обеспечивают насекомое витаминами и аминокислотами. Важнейший симбионт мухи цеце — бактерия Wigglesworthia — также населяет клетки бактериома мухи. Геном бактерии содержит гены, которые необходимы для синтезирования витаминов группы B, тогда как у мухи цеце имеются только лишь гены, необходимые для их транспортировки.
Кровеносная система
Кровеносная система насекомых незамкнутая. Гемолимфа только частично заключена в орган кровообращения — спинной сосуд, представляющий собой мышечную трубку. В остальном же она заполняет полость тела (миксоцель) и промежутки между органами, омывая их. Спинной сосуд разделён на 2 отдела: задний — сердце — и передний — аорту. Сердце разделено на камеры (от четырёх до десяти, обычно — 9), соответствующие брюшным сегментам. В эволюционно развитых группах имеется тенденция к уменьшению их числа. Каждая камера сердца имеет по два входных отверстия (устьица или остии) с клапанами, через которые гемолимфа поступает внутрь из полости тела. В отверстиях между камерами сердца также имеются клапаны, обеспечивающие движение гемолимфы в одном направлении. Задний конец сердца обычно закрыт. Аорта лишена камер и клапанов. В результате пульсаций камер сердца осуществляется движение гемолимфы в задне-переднем направлении (у некоторых насекомых наблюдается обратный ток гемолимфы). Частота сокращений сердца зависит от вида насекомого, его физического состояния, фазы развития и колеблется в пределах от 15 до 150 циклов в минуту. При диастоле гемолимфа входит в сердце через остии, а при систоле нагнетается вперёд, в аорту. Аорта открывается отверстием в полость головы, куда изливается гемолимфа. В полости тела насекомого кровообращение поддерживается двумя диафрагмами. Сокращение верхней диафрагмы увеличивает ёмкость околосердечной полости, куда устремляется гемолимфа в момент диастолы. Сокращение нижней диафрагмы способствует передвижению крови в полости тела спереди назад. Таким образом, совместная работа сердца и диафрагм осуществляет циркуляцию гемолимфы в организме насекомого: по спинному сосуду — сзади вперёд, в полости тела — спереди назад. Движение гемолимфы в придатках тела (усики, ноги, крылья) осуществляется дополнительными местными пульсирующими органами. Обычно это пульсирующие ампулы или подвижные мембраны (в ногах).
Нервная система

Схема строения нервной системы насекомых
Центральная нервная система (ЦНС) насекомых представляет собой цепь отдельных ганглиев, соединённых одиночными или парными коннективами. Самая передняя часть нервной системы, называемая мозгом, занимает дорсальное положение по отношению к пищеварительному каналу. Вся остальная часть ЦНС находится под кишечником. Мозг представляет собой три сросшихся ганглия и разделяется на три отдела: протоцеребрум, отвечающий за зрение и сложные поведенческие реакции, дейтоцеребрум — иннервирует антенны — и тритоцеребрум — иннервирует нижнюю губу, мышцы вокруг ротового отверстия, внутренние органы (высший центр иннервации внутренних органов). От мозга отходят две коннективы, огибающие кишечник с обеих сторон и сходящиеся на подглоточном ганглии, с которого начинается вентральная нервная цепочка. Её ганглии лежат под пищеварительным трактом. Подглоточный ганглий состоит из трёх сросшихся ганглиев и регулирует работу челюстей, гипофаринкса и слюнных желёз. Грудные ганглии развиты сильнее брюшных, так как регулируют работу конечностей и крыльев. Брюшные ганглии у прогрессивных форм имеют тенденцию к слиянию, а вся нервная цепочка - к укорочению. Так, у водомеркиHydrometra в груди имеется один слитный ганглий, а у высших двукрылых (Musca, Sarcophaga и др.) — два ганглия, подглоточный и общий грудной; в брюшке в обоих этих случаях ганглии вообще отсутствуют. Полный метаморфоз часто сопровождается концентрацией ЦНС — уменьшением числа ганглиев у имаго по сравнению с личинками.
У насекомых со сложным поведением (общественные насекомые) особенно сильно развит протоцеребрум, а в нём — увеличенная пара грибовидных тел. В них сосредоточены координирующие и высшие ассоциативные центры нервной системы и замыкаются условно-рефлекторные связи. Приобретённые навыки при их разрушении теряются (но безусловные рефлексы сохраняются). Наблюдается корреляция между сложностью поведения и степенью развития грибовидных тел. Например, среди медоносных пчёл грибовидные тела наиболее развиты у рабочих пчёл (выполняющих все работы), а наименее у трутней. В среднем размер грибовидных тел у медоносных пчёл равен 1⁄15 от размеров всего головного мозга, а у рабочих муравьёв рода формика они составляют почти половину всего мозга.
Вегетативная нервная система иннервирует внутренние органы. Она состоит из трёх отделов — краниального, туловищного и каудального. Краниальный отдел представлен стоматогастрической системой, которая иннервирует переднюю кишку и сердце. Стоматогастрическая система представляет собой соединённый с тритоцеребрумом фронтальный ганглий перед мозгом, соединённый с ним возвратным нервом затылочный ганглий за мозгом, а также кардиальные и прилежащие тела. Кардиальные тела осуществляют секрецию своих гормонов, а также накопление гормонов нейросекреторных клеток мозга. Основная функция прилежащих тел — синтез ювенильных гормонов. Туловищный отдел вегетативной нервной системы представлен непарным нервом, который считается аналогом симпатической нервной системы у позвоночных. В последнем брюшном ганглии непарный нерв образует две ветви, иннервирующие заднюю кишку и половые органы (каудальный отдел вегетативной нервной системы).
Пищеварительная система
Пищеварительная система насекомых состоит из кишечного тракта и пищеварительных желёз. У некоторых примитивных групп все три отдела кишечного канала имеют вид простой трубки, а у большинства насекомых каждый отдел дифференцирован на специализированные подотделы. У большинства насекомых имеются верхнечелюстные, нижнечелюстные и лабиальные слюнные железы. Наиболее развиты гроздевидные лабиальные слюнные железы, которые у гусениц превращаются в шёлкоотделительные органы, при этом функции слюнных желёз выполняются мандибулярными железами. В слюне содержатся различные ферменты: карбогидразы(амилаза, инвертаза), у фитофагов для расщепления углеводов; липазы и протеиназы для переваривания жиров и белков у хищников; пектиназа у тлей; гиалуронидазау клопов, разрушающая соединительные ткани; у кровососущих присутствуют антикоагулянты; ауксины, содействующие образованию галлов на растениях. У некоторых насекомых слюнные железы могут продуцировать шёлк, феромоны или аллотрофические вещества, выделяемые для питания других особей. Перепончатокрылые имеют также глоточные железы. Пчелиная матка с помощью феромонов мандибулярных желёз тормозит развитие яичников рабочих пчёл, у которых эти же железы выделяют маточное молочко для кормления личинок.
Кишечник разделён на три отдела: переднюю, среднюю и заднюю кишки, отделённые друг от друга клапанами. Кардиальный клапан отделяет переднюю кишку от задней, а пилорический — среднюю от задней. Стенки всех отделов кишечника образованы однослойным эпителием, который снаружи покрыт продольными и кольцевыми мышечными волокнами, обеспечивающими движение пищи в кишечнике. Вслед за глоткой расположен тонкий пищевод. Он может иметь дивертикулы изоб. Перед началом средней кишки расположен провентрикулус (мышечный желудок), образованный сильно развитыми кольцевыми мышцами и кутикулярными зубцами. Он обеспечивает дополнительное перетирание пищи и фильтрацию пищевых частиц, которые через кардиальный клапан передаются в среднюю кишку.
Основным органом переваривания и всасывания питательных элементов служит средняя кишка, где пищевые массы под действием ферментов расщепляются на простые органические соединения. Последние либо абсорбируются клетками эпителия стенок кишечника, либо диффундируют в гемолимфу. Строение средней кишки зависит от типа поглощаемой пищи. Задняя кишка разделена ректальным клапаном на тонкую и прямую кишки, и в ней происходят всасывание воды и формирование экскрементов. В тонкой кишке обитают симбионтные сапрофитные бактерии, простейшие и грибы, помогающие своим хозяевам (особенно сапрофагам, термитам и другим ксилофагам) в расщеплении целлюлозы до глюкозы. Некоторые микроорганизмы-симбионты насекомых могут синтезировать витамины, стерины и незаменимые аминокислоты. У некоторых насекомых пищеварительная система во взрослой стадии развития не функционирует. Например, у подёнок нет челюстей, а кишечник редуцирован; они не питаются и живут всего несколько часов или дней.
Выделительная система

Самец стрекозы Trithemis aurora
Мальпигиевы сосуды и задняя кишка служат основными органами выделения у преобладающего большинства насекомых. Наиболее примитивными органами выделения являются парные лабиальные органы щетинохвосток. Это гомологи метамерных органов кольчатых червей, а также антеннальных и максиллярных желёз ракообразных. Они состоят из начального мешочка, извитого лабиринта, выводного протока, открывающегося отверстием над основанием нижней губы.
У более совершенных групп насекомых основным органом выделения являются мальпигиевы сосуды. Они имеют вид трубочек, открывающихся на границе средней и задней кишок. Часто мальпигиевы сосуды свободно заканчиваются в гемолимфе. Их количество варьирует в различных группах от двух — восьми до 80—100 штук. В ряде случаев имеет место крептонефрия, когда кончики мальпигиевых сосудов прирастают к задней кишке (встречается у жуков, сетчатокрылых, гусениц и личинок пилильщиков), что уменьшает потерю влаги. Мальпигиевы сосуды и задняя кишка являются органами эмункториальной экскреции и освобождают организм насекомого от продуктов обмена веществ путём их выведения во внешнюю среду. В органах накопительной экскреции метаболиты накапливаются в клетках или тканях без непосредственного их выделения из организма. У насекомых такими органами являются жировое тело и перикардиальные клетки. Личинки перепончатокрылых, чьё развитие происходит в сотах, не выделяют экскременты до момента окукливания, и конечные продукты их метаболизма откладываются в жировом теле. Перикардиальные клетки располагаются вокруг аорты и сердечного сосуда, порой образуя скопления и в других участках тела. К дополнительным органам накопительной экскреции относятся эпидермальные клетки и их производные. У некоторых насекомых (тутовый шелкопряд, гладыши) выделительную функцию выполняют также некоторые клетки гиподермы, скидываемые во время линьки.
Формирование экскретов в мальпигиевых сосудах и задней кишке происходит в две фазы: фильтрация гемолимфы и обратное всасывание (реабсорбция) воды и полезных веществ в гемолимфу. Сформировавшиеся экскреты выводятся наружу вместе с экскрементами. Главным азотистым экскретом у наземных насекомых является мочевая кислота. Производные мочевой кислоты аллантоин и аллантоиновая кислота (является основным азотистым экскретом у некоторых чешуекрылых) также могут являться экскреторными продуктами. У водных насекомых и личинок мясных мух конечным продуктом азотистого обмена является аммиак.
Эндокринная система
Органы эндокринной системы насекомых синтезируют, накапливают и выделяют в гемолимфу различные гормоны, которые регулируют физиологические процессы. Основные эндокринные органы у насекомых представлены нейросекреторными клетками, ретроцеребральным комплексом, перисимпатическими органами ипроторакальными железами. Нейросекреторные клетки находятся в разных ганглиях ЦНС и вырабатывают нейрогормоны. Ретроцеребральный комплекс образован двумя парами мозговых желёз, называемых кардиальными, и прилежащими телами. Его функции заключаются в накоплении гормонов мозговых нейросекреторных клеток, а также продукции собственных гормонов, регулирующих обмен веществ, развитие, размножение и поведение насекомых. Основная функция прилежащих тел заключается в выработке ювенильных гормонов. Проторакальные железы производят экдизоны — гормоны, вызывающие линьку насекомых.
Половая система
Мужские половые железы насекомых представлены двумя семенниками, в которых происходит сперматогенез. Каждый из них образован фолликулами, исходное число которых равно семи. Вторичное увеличение количества фолликулов до 60—80 отмечается у прямокрылых и тараканов, в то время как у двукрылых и блох семенник состоит только из одного фолликула. Из семенников сперматозоиды попадают в систему семяпроводов, образованных парными семяпротоками и семяизвергательным каналом. Семяпротоки начинаются от семенников и заканчиваются семенными пузырьками. Они также сообщаются с мужскими придаточными железами, которые формируют сперматофор. Семяизвергательный канал выводит их наружу через мужские копулятивные органы. Обычно самцы насекомых имеют только одно половое отверстие и один копулятивный орган, расположенный на конце IX стернита брюшка. Но у самцов представителей отрядов подёнок и уховёрток имеется по два копулятивных органа. Структуры, обычно связанные с копулятивным органом, формируют мужские гениталии — эдеагус.
Женские половые органы насекомых представлены двумя яичниками и половыми путями. Яичники образованы отдельными яйцевыми трубками (овариолы), исходное количество которых равно семи. Однако у большинства насекомых наблюдается либо уменьшение, либо увеличение их количества. Например, почти все чешуекрылые имеют по 4 овариолы в яичнике. У матки медоносной пчелы количество овариол в среднем составляет 160 пар. Максимальное же число овариол — до 2000 в каждом яичнике — обнаружено у яйцекладущих самок термитов. Уменьшение числа овариол до одной-двух наблюдается у насекомых, которые перешли к паразитизму и живорождению. У всех насекомых, кроме подёнок, от яичников отходят два латеральных яйцевода, которые сливаются вместе, образуя непарный медиальный яйцевод. Обычно он сообщается с генитальной камерой. Для откладывания яиц и для введения копулятивного органа самца при спаривании служит наружное отверстие генитальной камеры. У большинства бабочек копулятивное и половое отверстия являются разделёнными и расположены на различных брюшных сегментах: VIII и IX. У живородящих насекомых яйца попадают из медиального яйцевода в расширение половых путей — матку, где в дальнейшем и происходит развитие эмбрионов.
Во время процесса копуляции самки получает от самцов сперму, сохраняющуюся в семяприёмнике и использующуюся для оплодотворения яиц при их откладывании. Придаточные железы женской половой системы у насекомых продуцируют клейкий секрет, который служит для приклеивания яиц к субстрату. У тараканов и богомоловон служит для образования оотеки. Стебельчатобрюхие перепончатокрылые характеризуются тем, что одна из придаточных желёз у них превращается в ядовитую железу, а другая продуцирует секрет для смазывания яйцеклада или жала.
Органы чувств
Насекомые, как и другие многоклеточные организмы, имеют множество различных рецепторов, или сенсилл, чувствительных к определённым раздражителям. Рецепторы насекомых очень разнообразны. У насекомых есть механорецепторы (слуховые рецепторы, проприоцепторы), фоторецепторы, терморецепторы, хеморецепторы. С их помощью насекомые улавливают энергию излучений в виде тепла и света, механические вибрации, включая широкий диапазон звуков, механическое давление, силу тяжести, концентрацию в воздухе водяных паров и летучих веществ, а также множество других факторов. Насекомые обладают развитым чувством обоняния и вкуса. Механорецепторами являются трихоидные сенсиллы, которые воспринимают тактильные стимулы. Некоторые сенсиллы могут улавливать малейшие колебания воздуха вокруг насекомого, а другие — сигнализируют о положении частей тела относительно друг друга. Воздушные рецепторы воспринимают скорость и направление потоков воздуха поблизости от насекомого и регулируют скорость полёта.
Зрение

Схема строения фасеточного глаза насекомого

Голова шершня. Кроме двух больших фасеточных глаз, хорошо видны 3 небольших глазка в области лба

Цветок водосбора голубого при обычном и УФ-освещении (внизу)
Зрение играет большую роль в жизни большинства насекомых. У них встречаются три типа органов зрения — фасеточные глаза, латеральные (стеммы) и дорсальные (оцеллии) глазки. У дневных и летающих форм обычно имеется 2 сложных глаза и 3 оцеллия. Стеммы имеются у личинок насекомых с полным превращением. Они располагаются по бокам головы в количестве 1—30 с каждой стороны. Дорсальные глазки (оцеллии) встречаются вместе с фасеточными глазами и функционируют в качестве дополнительных органов зрения. Оцеллии отмечены у имаго большинства насекомых (отсутствуют у многих бабочек и двукрылых, у рабочих муравьёв и слепых форм) и у некоторых личинок (веснянки, подёнки, стрекозы). Как правило, они имеются только у хорошо летающих насекомых. Обычно имеется 3 дорсальных глазка, расположенных в виде треугольника в лобно-теменной области головы. Их основная функция, вероятно, заключается в оценке освещённости и её изменений. Предполагается, что они также принимают участие в зрительной ориентации насекомых и реакциях фототаксиса.
Особенности зрения насекомых обусловлены фасеточным строением глаз, которые состоят из большого числа омматидиев. Наибольшее число омматидиев обнаружено у бабочек (12—17 тысяч) и стрекоз (10—28 тысяч). Светочувствительной единицей омматидия является ретинальная (зрительная) клетка. В основе фоторецепции насекомых лежит преобразование зрительного пигмента родопсина под воздействием кванта света в изомер метародопсин. Обратное его восстановление даёт возможность многократного повторения элементарных зрительных актов. Обычно в фоторецепторах обнаруживаются 2—3 зрительных пигмента, различающихся по своей спектральной чувствительности. Набор данных зрительных пигментов определяет также особенности цветового зрения насекомых. Зрительные образы в фасеточных глазах формируются из множества точечных изображений, создаваемых отдельными омматидиями. Фасеточные глаза лишены способности к аккомодации и не могут приспосабливаться к зрению на разных расстояниях. Поэтому насекомых можно назвать «крайне близорукими». Насекомые характеризуются обратно пропорциональной связью между расстоянием до рассматриваемого объекта и числом различимых их глазом деталей: чем ближе находится объект, тем больше деталей они видят. Насекомые способны оценивать форму предметов, но на небольших расстояниях от них для этого требуется, чтобы очертания объектов вмещались в поле зрения фасеточного глаза.
Цветовое зрение насекомых может быть дихроматическим (муравьи, жуки-бронзовки) или трихроматическим (пчелиные и некоторые бабочки). Как минимум один вид бабочек обладает тетрахроматическим зрением. Существуют насекомые, которые способны различать цвета только одной (верхней или нижней) половинкой фасеточного глаза (четырёхпятнистая стрекоза). Для некоторых насекомых видимая часть спектра сдвинута в коротковолновую сторону. Например, пчёлы и муравьи не видят красного цвета (650—700 нм), но различают часть ультрафиолетового спектра (300—400 нм). Пчёлы и другие насекомые-опылители могут увидеть на цветках ультрафиолетовые рисунки, скрытые от зрения человека. Аналогично бабочки способны различать элементы окраски крыльев, видимые только в ультрафиолетовом излучении.
У многих насекомых глаза развиты слабо или отсутствуют. Это виды, живущие в подстилке и почве (многие термиты,бессяжковые и др.), многие пещерные виды, некоторые паразиты (напр., вши рода Haematopinus).
Слух
Восприятие звуков, передающихся через твёрдый субстрат, осуществляется у насекомых виброрецепторами, находящимися в голенях ног вблизи их сочленения с бедром. Многие насекомые обладают высокой чувствительностью к сотрясениям субстрата, на котором они находятся. Восприятие звуков через воздух или воду осуществляется фонорецепторами. Двукрылые воспринимают звуки при помощи джонстоновых органов. Наиболее сложными слуховыми органами насекомых являются тимпанальные органы. Количество сенсилл, входящих в состав одного тимпанального органа, варьирует от 3 (некоторые бабочки) до 70 (саранчовые) и даже до 1500 (у певчих цикад). У кузнечиков, сверчков и медведок тимпанальные органы находятся в голенях передних ног, у саранчовых — по бокам первого брюшного сегмента. Слуховые органы певчих цикад располагаются у основания брюшка в близости от звукопроизводящего аппарата. Слуховые органы ночных бабочек находятся в последнем грудном сегменте или в одном из двух передних сегментов брюшка и могут воспринимать ультразвуки, издаваемые летучими мышами.Медоносные пчёлы издают звуки, заставляя вибрировать часть торакса путём частых мышечных сокращений. Звук усиливается крыловыми пластинами. В отличие от многих насекомых пчёлы способны издавать звуки разной высоты и тембров, что позволяет им передавать информацию посредством разных характеристик звука.
Обоняние и вкус
Насекомые обладают развитым обонятельным аппаратом. Восприятие запахов осуществляется благодаря хеморецепторам — обонятельным сенсиллам, расположенным на усиках, а иногда и на околоротовых придатках. На уровне хеморецепторов происходит первичное разделение обонятельных раздражителей благодаря наличию двух типов рецепторных нейронов. Нейроны-генералисты распознают очень широкий набор химических соединений, но при этом обладают низкой чувствительностью к запахам. Нейроны-специалисты реагируют только на одно или несколько родственных химических соединений. Они обеспечивают восприятие пахучих веществ, запускающих определённые поведенческие реакции (половые феромоны, пищевые аттрактанты и репелленты, углекислый газ). У самцов тутового шелкопряда обонятельные сенсиллы достигают теоретически возможного предела чувствительности: для возбуждения нейрона-специалиста достаточно всего лишь одной молекулы феромона самки. В своих опытах Ж. А. Фабр определил, что самцы грушевой павлиноглазки могут обнаруживать самок по феромонам на расстоянии до 10 км.
Контактные хеморецепторы образуют периферический отдел вкусового анализатора насекомых и позволяют им оценивают пригодность субстрата для питания или яйцекладки. Эти рецепторы располагаются на ротовых частях, кончиках лапок, антеннах и яйцекладе. Большинство насекомых способны распознавать растворы солей, глюкозы, сахарозы и других углеводов, а также воду. Хеморецепторы насекомых редко реагируют на искусственные вещества, имитирующие сладкий или горький вкус, в отличие от хеморецепторов позвоночных. Например, сахарин не воспринимается насекомыми как сладкое вещество.