|
|
Развитие и размножение
Размножение
Двуполое размножение

Спаривающаяся пара двух цветовых вариаций тополёвых бражников (Laothoe populi)
Размножение у насекомых преимущественно осуществляется двуполым путём, который включает: осеменение, оплодотворение и откладывание яиц или отрождение личинок. Типичное наружно-внутреннее осеменение среди насекомых характерно для подкласса первичнобескрылых (Apterygota). У крылатых насекомых (Pterygota) типичное наружно-внутреннее осеменение не встречается, а перенос спермы у них всегда сопровождается процессом спаривания. У представителей многих отрядов крылатых насекомых отмечается внутреннее сперматофорное осеменение, при котором сперматофоры прямо вводятся в половые пути самки либо подвешиваются к ним. При внутреннем осеменении сперматофор защищает семенную жидкость от высыхания и обеспечивает её питательными веществами. Оболочка сперматофора растворяется в половых путях самки, сперматозоиды активируются и мигрируют в семяприёмник. У насекомых, чьё спаривание протекает без сперматофоров, при копуляции семенная жидкость свободно вводится самцом в половые пути самки.
Другие способы размножения

Живорождение у тлей
Ряд насекомых, обычно размножающихся обоеполым путём, характеризуются способностью к спонтанному партеногенезу, при котором яйца развиваются без оплодотворения. Данный тип партеногенеза известен у тутового шелкопряда. В то же время популяционный партеногенез встречается во всех отрядах насекомых, но чаще у сеноедов. Партеногенетические виды и расы насекомых обычно проживают в условиях пониженной активности и недостаточной численности полов, которые затрудняют возможность встречи особей для спаривания. Например, европейский палочник (Bacillus rossius) повсеместно размножается обоеполым путём, но на севере ареала переходит к партеногенетическому размножению. У многих видов сеноедов существуют популяции двух типов: обоеполые и партеногенетические (совсем не имеющие самцов). При этом самки из партеногенетических популяций теряют способность спариваться с самцами. Циклический партеногенез проявляется чередованием партеногенетических и обоеполых поколений. Его примерами служат тли и галлицы, у которых партеногенез сочетается с живорождением. Летом партеногенетические самки-девственницы рождают личинок, а появляющиеся к осени оплодотворённые самки обоеполого поколения откладывают зимующие яйца. У галлиц из подсемейства Lestremiinae циклический партеногенез сочетается не только с живорождением, но и с неотенией. Избирательный партеногенез отмечается у перепончатокрылых: из оплодотворённых яиц развиваются самки, а из не оплодотворённых — самцы. Своеобразна и сложна биология размножения жука микромальтуса, у которого известны три личиночных возраста. Личинки первого возраста очень подвижны, но не питаются. После линьки они превращаются в активно питающихся безногих личинок 2-го возраста. Они, в свою очередь, превращаются в личинок 3-го возраста, которые не питаются, но способны к размножению и рождают множество личинок 1-го возраста. Известны у данного вида иимаго, развивающиеся из личинок второго возраста. У микромальтуса имеет место единственный среди насекомых случай сочетания гиперметаморфоза с размножением личинок — педогенезом. Среди насекомых бесполое размножение отмечено только на эмбриональной стадии у веерокрылых и у некоторых эндопаразитических перепончатокрылых. Этот способ размножения, получивший название полиэмбрионии, заключается в том, что из одного яйца развивается не один зародыш, а несколько.
Брачные ритуалы

Два дерущихся самца жука-оленя

Самец пчелы Euglossa с расширенными голенями задних ног, концентрирующими летучие соединения для привлечения самок
Многие группы насекомых обладают развитыми брачными ритуалами, которые предшествуют самому процессу спаривания. Сложные формы ухаживания, как то брачные полёты и «танцы», отмечаются у многих бабочек. Например, самцы хвостатки летят под самкой, а затем разворачиваются в другую сторону, касаясь крыльями усиков самки; у лимонницы самец летит позади самки, сохраняя постоянную дистанцию. У других видов бабочек самка может сидеть на растении, а самец совершает «танцующий» полёт прямо над ней, выделяя феромоны, стимулирующие её к спариванию.
Самцы некоторых насекомых, например жуков-рогачей, сражаются за самок на «брачных турнирах». Самцы австралийской одиночной пчелы Доусона часто убивают друг друга в драках за самку. При этом отмечается диморфизм самцов, например, у калоеда-быка, Podischnus agenor и др. Обладая глазами на длинных «стебельках», самцыстебельчатоглазых мух устраивают драки между собой за целые «гаремы» самок, собирающихся группами на ночлег. Самцы ряда скорпионниц, например Hylobittacus apicalis, приносят в подарок самке пойманное насекомое, и чем больше добыча, тем выше вероятность начала спаривания. У толкунчиков самец при ухаживании также преподносит самке подарок: лепесток цветка или добычу — другое насекомое. Самцы орхидных пчёл имеют уникальные видоизменённые широкие задние ноги, используемые для сбора и хранения различных летучих соединений (чаще эфиров), собираемых ими на различных цветках и служащих для привлечения самок.
У некоторых видов насекомых самцы могут оставаться с самкой после спаривания и охранять её от других самцов, пока она не отложит яйца. Самцы бабочки Heliconius charithonia начинают спариваться с самкой ещё до окончания её полного выхода из куколки. Данное явление в англоязычной литературе получило название «изнасилование куколки» (англ. pupal rape).
Жизненный цикл
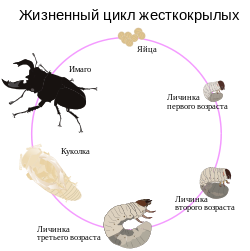
Схема полного метаморфоза насекомых на примере жуков-рогачей
Схема неполного метаморфоза насекомых на примере саранчовых
Весь жизненный цикл, называемый генерацией или поколением, протекает у разных видов в течение неодинакового времени. Многие виды имеют короткую продолжительность генерации. Самый короткий половой жизненный цикл (от откладывания яйца до выхода имаго) обнаружен у следующих видов насекомых: комар Psorophora confinnis менее 1 недели; муха Haematobia irritans — 9 дней. Некоторые виды способны давать в год до 10—15 поколений (ряд тлей, некоторые тропические бабочки) и более. У других одна генерация продолжается много лет (так, у чёрного таракана развитие длится 4 года, а у семнадцатилетней цикады 17 лет). Другой существенной стороной жизненного цикла видов является время продолжительности тех или иных фаз развития. Так, зимовка видов, обитающих в умеренном поясе, может проходить на стадии яйца, личинки, куколки либо имаго; соответственно этому меняется время длительности других фаз на протяжении вегетационного периода. Виды могут отличаться друг от друга не только количеством генераций в году, но и временем протекания отдельных стадий развития, то есть годичным циклом. Иногда годичный цикл осложняется задержкой развития —диапаузой, сопровождающейся понижением обмена веществ и прекращением питания. Диапауза контролируется как внешними (температура, длина светового дня и др.), так и внутренними (гормональными) факторами, являясь одной из форм приспособления насекомых к жизни в областях с резкими сезонными изменениями климата. Многие диапаузирующие насекомые при вынужденном развитии без диапаузы (например, в лабораторных условиях) испытывают нарушения онтогенеза или погибают.
Большинство насекомых первично откладывает яйца, но некоторые виды способны к отрождению личинок. Факультативное живорождение, когда самки могут откладывать яйца и отрождать личинок, отмечено у некоторых жуков-листоедов и стафилидов. Облигатное живорождение сопровождается возникновением матки, в которой происходит развитие эмбрионов. При истинном живорождении отмечаются более тесные взаимоотношения между материнским организмом и развивающимися в нём эмбрионами. При этом эмбрионы снабжаются от самки «молочком», содержащим необходимые питательные вещества. Способность кяйцеживорождению встречается у многих групп насекомых. Особо следует упомянуть о живорождении тлей и галлиц, у которых оно является сцепленным с циклическим партеногенезом. Истинное живорождение отмечено также у тараканов рода Diploptera и некоторых мух (Cyclorrhapha), отрождающих вполне зрелых личинок, которые не приступают к питанию и окукливаются.
Яйца насекомых могут быть различной формы, с прочной хитиновой оболочкой, иногда имеющей сложную скульптуру. Их размеры от долей миллиметра до 16,5 мм в длину при диаметре 3 мм (у древесной пчелы Xylocopa auripennis). Количество откладываемых яиц от 1 (у ряда тлей) до десятков тысяч и более. Бабочка Trictena atripalpis из Австралии отложила 29 100 яиц, а при вскрытии в ней нашлось ещё около 15 тысяч яиц. Мирмекофильные наездники Kapala terminalis и Stilbula cynipiformis откладывают до 15 000 яиц. Матка термита Termes bellicosus откладывает около 30 тысяч яиц в сутки на протяжении примерно 10 лет своей жизни (в сумме боле 100 млн яиц). Муравьиная матка Eciton burchellii кладёт до 120 тысяч яиц каждые 36 дней, а матка кочевого муравья Dorylus wilverthiможет отложить до 4 млн яиц за один месяц. Самоё маленькое потомство приносят мухи-кровососки вида Hippobosca variegata (4 личинки) и муха цеце Glossina palpalis (6—12 личинок).
Способы откладывания яиц разнообразны. Многие насекомые просто разбрасывают яйца, периодически или партиями выделяя их. Другие — прикрепляют их к субстрату. В ряде случаев кладки яиц покрываются пенистыми, слизистыми или прочими выделениями.
После завершения развития в яйце (эмбриогенеза) происходит вылупление. С этого момента насекомое вступает во второй этап своего развития — фазу личинки, с которой и начинается послезародышевый период. У насекомых прослеживается тенденция к разделению жизненных функций между личинками и имаго. Во время фазы личинки происходит рост и накопление питательных веществ, а в фазе имаго — размножение и расселение.
Полный и неполный метаморфоз

Гусеница бабочки Calliteara pudibunda
Высших, или крылатых, насекомых разделяют на насекомых с полным и неполным превращением. Преобладающее большинство видов насекомых с полным метаморфозом проходят в своём развитии четыре фазы: яйцо, личинка, куколка и имаго. Однако имеются некоторые исключения (например, нарывники) — наблюдается явление гиперметаморфоза с пятью — шестью и более фазами, которые представлены в виде дополнительных, особых личиночных стадий, непохожих на основную личинку и имаго. У нарывников рода Meloe и Mylabris данная дополнительная стадия называется «триунгулин». Другим исключением является неотения, представляющая собой способность достигать половой зрелости и размножаться на стадии личинки, например, у самок жуков рода Platerodrilus. При этом в развитии самок исключаются сразу две фазы: куколка и имаго (однако данное явление не исключает нормального развития самцов).
Неполное превращение характеризуется прохождением насекомым трёх фаз развития (яйцо, личинка и имаго). При этом у насекомых с неполным превращением личинки внешне похожи на взрослых насекомых и отличаются от них меньшими размерами, отсутствием или зачаточными наружными половыми придатками и крыльями.
У насекомых с полным превращением личинки обычно имеют червеобразную форму тела, и только имаго обладают всеми особенностями, характерными для отряда. У большинства видов насекомых с неполным превращением имаго и личинки ведут сходный образ жизни. Личинки насекомых весьма разнообразны, но могут быть сведены к двум основным типам — сходным со взрослой особью и несходным; первые называются нимфами, вторые — истинными личинками. Нимфы сходны со взрослой стадией как морфологически, так и биологически: они живут обычно в той же среде, что и имаго, и сходно питаются. Истинные личинки резко отличаются от взрослых особей внешним видом, средой обитания и пищей. Именно таким личинкам для превращения во взрослое насекомое требуется стадия куколки. У некоторых водных насекомых превращение личинок в имаго проходит через промежуточную стадию — субимаго. Оно имеет внешние особенности взрослых насекомых, но не развито в половом отношении.
Рост и развитие личинок сопровождаются периодическими линьками — сбрасыванием кожных покровов, благодаря чему происходят рост тела и его наружные изменения. Количество линек личинки неодинаково у разных насекомых и варьирует от трёх (мухи, многие жуки) или четырёх — пяти (бабочки, прямокрылые, клопы и др.) до 25—30 у подёнок и веснянок. С каждой новой линькой личинка вступает в следующую стадию, или возраст. Количество линек соответствует числу личиночных возрастов.
В конце концов личинки переходят во взрослое насекомое — имаго. У насекомых с неполным превращением это происходит во время последней линьки личинки, а с полным превращением — посредством особой фазы развития, называемой куколкой. Обычно куколки неподвижны, но иногда способны к передвижению благодаря ритмичным изгибаниям тела (у верблюдок, некоторых мух). Иногда предкуколочное состояние покоя обозначается как особая стадия — предкуколка. Куколки разных насекомых весьма отличаются друг от друга особенностями строения. Нередко перед окукливанием личинки окружают себя коконом, который чаще всего сооружается из шёлка. Часто шёлк служит основой для скрепления частиц субстрата, из которого формируется кокон. Многие личинки жуков и гусеницы бабочек не сооружают себе кокон, но изготовляют из окружающего субстрата и своих экскрементов округлую полость, образуя т. н. «колыбельку». Имаго является заключительной стадией жизненного цикла.
Особую форму метаморфоза, проходимую совместно с основным, представляет собой так называемая фазовая изменчивость, открытая первоначально у саранчовых, но затем обнаруженная и у остальных прямокрылых, а также у некоторых палочников, бабочек и других насекомых. Сущность её заключается в том, что при возникновении скученности особей изменяются морфологические и биолого-физиологические свойства вида.
В стадии имаго насекомые не совершают линек и не способны к росту. Исключение составляют лишь подёнки и т. д.
Численность того или иного вида насекомых не остаётся постоянной. Из года в год, в зависимости от природных условий, количество особей одного вида может меняться в десятки, сотни и тысячи раз. Периодически возникающие массовые размножения называются «вспышками численности». После подобных вспышек, как правило, следует снижение численности, и некоторое время вид может встречаться редко, после чего происходит восстановление обычной численности.
Продолжительность жизни

Цикады Magicicada septendecula живут в стадии личинок под землёй на протяжении 17 лет
На взрослой стадии развития насекомые живут от нескольких часов до десятков лет. Например, имаго подёнок живут от нескольких часов до нескольких дней. Мало живут также имаго веснянок, ручейников. В то же время на стадии личинок подёнки живут 2—3 года. Самая короткая продолжительность жизни взрослой особи отмечена у подёнок Dolania americana. Самка этого вида живёт 5 минут после последней личиночной линьки (за это время она спаривается, откладывает яйца). Продолжительность жизни большинства бабочек (за редким исключением) составляет до двух — трёх недель. Более двух лет требуется для развития стрекоз и некоторых жуков. Например, 2 года длится развитие хлебного жука-кузьки, 3—4 года — у жуков-щелкунов и майского жука, ещё больше этот срок может быть у златок, дровосеков, чернотелок. Например, жук-олень на стадии имаго живёт 1—2 месяца, но его личинка развивается преимущественно в древесине дуба на протяжении четырёх — шести лет. Наоборот, продолжительность жизни имаго чернотелки Polposipus herculeanus с Сейшельских островов может составлять до восьми лет. Рекордсменом среди бабочек стала живущая в суровых условиях Гренландии бабочка Gynaephora groenlandica, известная тем, что жизненный цикл её гусеницы может длиться до семи и даже до 14 лет (большую часть жизни они проводят в диапаузе). Имеет место также различная продолжительность жизни самок и самцов. Например, самка жука-оленя живёт в 2 раза дольше самца, примерно то же соотношение наблюдается у комнатной мухи.
Североамериканские певчие периодические цикады Magicicada живут в стадии личинок под землёй 13 и 17 лет в зависимости от вида. Однако рекорды в этой области, как правило, принадлежат общественным насекомым, чьи крупные гнёзда (муравейники и термитники) могут наблюдаться живыми многие десятилетия. Муравьиные матки живут до 20 лет, а царицы термитов — до 25. В то же время рабочие особи тех же видов живут от нескольких месяцев до трёх лет, а самцы — всего несколько недель.
Косвенные расчёты показывают ещё большую продолжительность жизни диапаузирующих преимагинальных стадий развития насекомых. Например, известны данные (в том числе занесённые в Книгу рекордов Гиннесса), указывающие на то, что возраст личинок жуков-златок может быть от 26 лет до 51 года. Однако они основаны только на возрасте мёртвой древесины, из которой зафиксирован выход златок Buprestis aurulenta.
Забота о потомстве
Насекомые характеризуются развитым инстинктом заботы о потомстве. Это проявляется прежде всего в заготовке запасов пищи для будущих личинок либо в уходе за яйцами (или даже за личинками). Наиболее примитивное проявление заключается в обеспечении безопасности откладываемых яиц путём сбережения их в естественных убежищах, например, трещинах почвы или древесной коры, зарывания их в землю и т. п. Некоторые жуки прикрывают своё потомство собственным телом — южноамериканские жуки-щитоноски из рода Omaspides обладают расширенными боковыми краями надкрылий, под которыми укрываются до 20 молодых личинок. Некоторые клопы, например Phyllomorpha laciniata, откладывают яйца на спинную сторону тела самца, где они находятся до выхода личинок. Ряд долгоносиков, например долгоносики-плодожилы (Balaninus), проделывают отверстия в плодах разных деревьев и откладывают в них свои яйца. Листоеды из рода Phyllodecta изготавливают защитную капсулу для яиц из собственных экскрементов, ряд ночных бабочек покрывают отложенные яйца волосками со своего тела, тропические жуки-щитоноски рода Aspidomorpha сооружают настоящие оотеки. Ещё одним способом является изготовление из листьев т. н. «сигар» (или кульков-фунтиков) трубковёртами. Заготовка корма для личинок достигает сложности у жуков-навозников и включает в себя как рытьё норок под кучами помёта, так и транспортировку его (после скатывания в шары) на большие расстояния в сложную систему подземных камер и ходов. Мертвоеды (Silphidae), в частности жуки-могильщики, обеспечивают своих личинок пищей путём зарывания в землю трупов позвоночных животных.
Самки обитающего в Японии древесного клопа Parastrachia japonensis ежедневно приносят в гнездо к своим личинкам плоды дерева Schoepfia jasminodora. Охраняют свои яйца и молодых личинок уховёртки. При этом самка для защиты яиц от плесени постоянно их облизывает. Ещё более сложное поведение обнаружено у сверчка Anurogryllus (Gryllidae), обитающего в Северной Америке. Самка этого вида, кроме охраны отложенных ею яиц, ещё и регулярно приносит молодым личинкам пищу, а также кормит их мелкими и бесплодными кормовыми яйцами. Повзрослевшие личинки старших возрастов в дальнейшем покидают гнездо и живут отдельно от матери.
Признаки эусоциальности обнаружены у некоторых колониальных тлей, отдельных видов трипсов. У жуков-плоскоходов Austroplatypus incompertus, кроме заботы о потомстве, обнаружены разделение на плодовитые и бесплодные касты и перекрывание нескольких поколений. Жуки рода Pselaphacus из семейства грибовиков (Erotylidae) держатся группами и ведут своих личинок к грибам, которые те потом будут поедать. Представители семейства пассалидов (Passalidae) отличаются высокоразвитым инстинктом заботы о потомстве. Обитая в гнилой древесине, эти жуки часто встречаются в ней целыми скоплениями, образуя социальные группы. Самцы и самки, живя парами, совместно выкармливают своё потомство пережёванной древесиной, прошедшей ферментацию. Они также охраняют куколок и недавно вышедших из них имаго.
Наибольшее разнообразие форм заботы о потомстве наблюдается среди жалящих перепончатокрылых, таких как осы и пчёлы. При этом различают разные стадии перехода от одиночного образа жизни к общественному (пресоциальность, субсоциальность, полусоциальность, парасоциальность и квазисоциальность). Высшая стадия заботы о потомстве (эусоциальность муравьёв, медоносных пчёл и т. д.) встречается у общественных насекомых.
Целый ряд одиночных ос охотятся на других насекомых или пауков, которых они парализуют жалом, после чего переносят в гнездо, где кормят ими своих личинок. Например, аммофилы охотятся на гусениц бабочек, дорожные осы (например, Pepsis, Agenioideus) — на пауков, пчелиный волк — на пчёл, сфексы — на саранчовых и кузнечиков, Sphecius — на цикад, Ampulex compressa — на тараканов, мелкие осы-пемфредонины — на тлей, трипсов и коллембол. Одноразовое снабжение пищей личиночных камер и их запечатывание после откладывания яйца считаются примитивными. Более эволюционно продвинутые осы выкармливают своё потомство постепенно, периодически принося ему очередные порции свежего корма в виде парализованных жертв. Например, одиночные осы-бембексы ежедневно приносят своим личинкам по несколько убитых мух (Tabanidae, Syrphidae).

Скарабей заготавливает пищу для будущих личинок

Дорожная оса Auplopus carbonarius с парализованным ею пауком

Трубковёрт Apoderus coryli за сооружением «сигары» из листа

Роющая оса Ammophila sabulosa, переносящая гусеницу

Самка уховёртки в гнезде, охраняющая свой приплод

Лист, свёрнутый трубковёртом Apoderus erythrogaster
Гинандроморфизм
Во время развития насекомых с полным превращением возможны генетические изменения, приводящие к появлению гинандроморфных особей — полусамцов-полусамок, у которых одна сторона тела полностью мужская, а другая — женская (т. н. билатеральные гинандроморфы). Частота появления гинандроморфов составляет примерно один случай на 90 000. Основой для появления гинандроморфа обычно является образование двуядерных ооцитов, у которых одно из ядеробразуется, например, из ядра полярного тельца.
Межвидовое скрещивание
Межвидовое скрещивание особей различных видов насекомых наблюдается как в природе, так и при культивировании человеком (содержании в неволе) у целого ряда видов, преимущественно чешуекрылых, ряда жесткокрылых и некоторых перепончатокрылых. Гибриды насекомых могут быть внутривидовыми (при скрещивании различных подвидов), внутриродовыми (при скрещивании видов, принадлежащих одному роду) или межродовыми (при скрещивании видов, относящихся к разным родам). Как правило, межвидовые гибриды не способны к размножению.