|
|
Эволюция и систематика
Происхождение
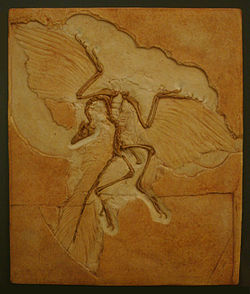
Археоптерикс. Берлинский экземпляр
Происхождение птиц долгое время оставалось предметом оживленных дискуссий. За обозримый период времени было выдвинуто несколько научных версий происхождения и родственных связей птиц и возникновения у них полёта, причём в течение более чем ста лет они были чисто гипотетическими.
Впервые теория эволюции птиц от рептилий возникла после открытия в 1860 году в Германии окаменелых останковархеоптерикса — животного, жившего около 150 млн лет назад в верхней юре. Он обладал характеристиками типичного пресмыкающегося — особым строением таза и рёбер, зубами, когтистыми лапами и длинным, как у ящерицы, хвостом. При этом окаменелости имели хорошо сохранившиеся отпечатки маховых крыльев, аналогичные таковым у современных птиц. На протяжении многих десятилетий история развития птиц рассматривалась как эволюция группы, развивавшейся от археоптерикса.
Именно на его изучении и основывались все первые гипотезы и теории о происхождении и родственных связях птиц: теория древесного («с деревьев вниз», Marsh, 1877) и теория бегательного («с земли вверх», Williston, 1879) возникновения полёта у птиц. Соответственно этим представлениям трактовалось и происхождение самих птиц — от триасовых текодонтов(архозавроморф) при древесной теории или от юрских бегающих тероподных динозавров при наземной теории.
В настоящее время археоптерикса уже не рассматривают в качестве общего предка всех современных птиц. Однако, он, вероятно, имеет близкое родство с их настоящим предком. Точное положение археоптерикса в эволюционном дереве определить трудно. Согласно кладистическому анализу китайских палеонтологов Xu et al. 2011 года, археоптерикс может представлять собой (вместе с видом Xiaotingia zhengi) параллельную тупиковую ветвь на общем стволе динозавров. Однако, более тщательный филогенетический анализ не подтвердил размещение археоптерикса среди дейнонихозавров и поэтому его продолжают считать древнейшей и наиболее примитивной птицей (в составе группы Avialae). Тем не менее, были обнаружены более древние ископаемые останки, которые также могут быть отнесены к кладе Avialae, но на данный момент они рассматриваются как динозавры:Anchiornis, Xiaotingia и Aurornis.
Динозавры и птицы

Голотип Microraptor gui с перьями
Гипотеза о происхождении птиц от динозавров впервые была выдвинута в 1868 году Томасом Гексли. Она основывалась на сравнении строения археоптерикса и маленького хищного динозавра рода Compsognathus.
С течением времени, и особенно в последние десятилетия, в связи с открытием многих видов древних птиц и оперённых динозавров-теропод, большинство исследователей стали склоняться к теории, согласно которой птицы произошли от динозавров-тероподов из группы манирапторов.

Реконструкция летающего хищного динозавра чанъюйраптора, покрытого перьями
По мере обнаружения всё большего количество ископаемых останков не летающих тероподов, так или иначе имеющих родство с птицами, чёткая граница между «птицами» и «не-птицами» становится размытой, и провести различие между данными группами всё сложнее. Если ранее одним из определяющих признаков птиц считалось наличие перьевого покрова, то ряд открытий конца XX — начала XXI веков в провинцииЛяонин на северо-востоке Китая показывает, что многие мелкие тероподы, например синозавроптерикс и синорнитозавр, имели перья. Эволюционное происхождение перьев можно проследить начиная с карнотавра, покрытого многочисленными бугорками, похожими на фолликулы. Следующий этап виден у синозавроптерикса и Dilong paradoxus, покрытых волокнистым пухом. Наконец у каудиптерикса, протархеоптерикса, синорнитозавра, микрораптора и Changyuraptor yangi уже имелись настоящие перья.
| ||||||||||||||||||||||||||||||
| Кладограмма филогении птиц согласно Godefroit et al., 2013. |
Птиц роднит с динозаврами множество общих черт в строении скелета. В последние годы среди палеонтологов достигнут консенсус, что ближайшими родственниками птиц являются дейнонихозавры — инфраотряд, включающий в себя семействадромеозавров и троодонтид. Эти три группы были объединены в единую, получившую название Paraves.
Центральное место среди дромеозавров занимает «четырёхкрылый» микрораптор, который был способен летать либо планировать. Тот факт, что дейнонихозавры преимущественно были маленького размера, дал основание предполагать, что предок птиц вёл древесный образ жизни и был способен к планированию.
Среди возможных ближайших родственников птиц также рассматривается авимим — мелкий хищный динозавр из мелового периода.
Альтернативные версии происхождения

Реконструкция Longisquama insignis
Наряду с теорией происхождения от динозавров существует гипотеза, согласно которой птицы могли эволюционировать отархозавроморфов независимо от линии развития динозавров, в поздней перми или раннем триасе. Раньше таковыми рассматривались мелкие и более примитивные псевдозухии из группы текодонтов, теперь в качестве возможных предков преимущественно рассматривают орнитозухид. Впервые данная теория была высказана в 1913—1914 годах Робертом Брумоми Фридрихом Хюне, позднее защищалась Герхардом Хейельманом, и оставалась наиболее популярной вплоть до конца двадцатого столетия. Тем не менее, некоторые учёные, среди которых американские палеонтологи Ларри Мартин (англ.)русск. и Алан Федучия (англ.)русск., продолжают настаивать, что птицы произошли от более ранних архозавров типа лонгисквамы. Они утверждают, что сходство между птицами и манирапторами является результатом конвергентной эволюции. В конце 1990-х годов Федучия и Мартин выдвинули модифицированную версию своей гипотезы, согласно которой манирапторы являются нелетающими потомками ранних птиц. Данная гипотеза оспаривается большинством палеонтологов. В июле 2014 года в продолжение своей гипотезы Алан Федучия в соавторстве опубликовал исследование, согласно которому Scansoriopteryxрассматривался ими как древняя птица — потомок архозавров. По мнению авторов публикации, наличие у него архаичных черт и анатомических особенностей, характерных для птиц, и перьев на конечностях свидетельствуют о возникновении полёта «сверху вниз», при котором архозавры обзавелись перьями. Также ими утверждается, что Scansoriopteryx и все птицы произошли от архозавров, таких как склеромохлус.Филогенетические исследования никогда не подтверждали данную альтернативную интерпретацию, и прежние аргументы авторов публикации ранее уже неоднократно были подвергнуты критике как ненаучные.
Ещё одну версию происхождения птиц отстаивал российский палеонтолог Евгений Курочкин. По его мнению, предком настоящих птиц мог быть протоавис, который на 75 млн лет древнее археоптерикса. Критики данной теории утверждают, что останки протоависа плохо сохранились и могут быть химерой.
В 1977 году палеонтолог Алик Уолкер, после изучения скелета триасового южноафриканского текодонта-крокодиломорфа сфенозуха (Sphenosuchus), предложил гипотезу о происхождении птиц от ранних, позднетриасовых крокодилов. Она базировалась на большом количестве общих черт строения черепа ранних крокодилов и птиц. Аргументы Уолкера неоднократно критиковались, и сам автор впоследствии признал гипотезу несостоятельной.
Помимо описанных выше основных теорий происхождения птиц, были выдвинуты и многие другие. Однако, все эти гипотезы не принимаются сегодня всерьёз. Это теория Фогта о родстве птиц и ящериц (1879), теория о связи птиц с птерозаврами (Оуэн, 1875; Сили, 1881), теория о происхождении птиц от эозухий (Г. Дементьев, 1940), теория о родстве птиц и птицетазовых динозавров (Galton, 1970).
Ранняя эволюция птиц
| Филогения по Chiappe (2007) | |||||||||||||||||||||||||||
|

Конфуциусорнис — примитивная ископаемая птица из раннего мелового периода. Китай
Наиболее ранние представители клады Avialae — птицеподобные динозавры Anchiornis huxleyi, Xiaotingia zhengi,Aurornis xui, известные по окаменелостям из геологической формации Тяоцзишань в Китае, датируемой концомюрского периода (около 160 млн лет назад). Многие из этих ранних форм обладают необычными анатомическими особенностями, которые в дальнейшем были утеряны птицами в ходе эволюции.
Диверсификация, или эволюционное разделение на множество разнообразных форм, началось в меловом периоде. Многие из этих форм, в том числе и предки современных веерохвостых птиц, на первоначальном этапе сохраняли примитивные признаки от динозавров — крылья с когтями и зубы, которые впоследствии были редуцированы. Джехолорнис и другие древнейшие птицы обладали длинным хвостовым отделом позвоночника. У представителей других, более развитых групп, например клады пигостилевых, 4—6 задних хвостовых позвонков были слиты воедино, образовав пигостиль.
Первой широко распространённой и разнообразной группой короткохвостых птиц стали энанциорнисовые, или «противоположные птицы», названные так потому, что строение их плечевых костей было противоположно строению таковых у современных птиц. Примитивные энанциорнисовые не связаны общим происхождением с более прогрессивными веерохвостыми птицами и представляют отдельную ветвь эволюции. Внешне похожие на современных птиц и развивавшиеся вместе с ними в меловом периоде, энанциорнисы заняли различныеэкологические ниши: от обитателей песчаных морских побережий и охотников за рыбой до ведущих древесный образ жизни и питающихся семенами. Более развитые ихтиорнисы, несколько напоминающие современных чаек, обитали на морских просторах и также питались рыбой. Морские птицы из отряда гесперорнисообразных из мезозоянастолько приспособились к охоте за рыбой, что в ходе эволюции утратили способность к полёту и полностью перешли к водному образу жизни. Несмотря на такую узкую специализацию, они являются одними из родственников современных птиц.
Эволюция современных птиц

Rapaxavis pani — ископаемый вид энанциорнисовых птиц
Обнаруженные на острове Вега у берегов Западной Антарктиды останки древней птицы вегавис, показали, что по крайне мере одна эволюционная линия веерохвостых птиц получила развитие в начале мелового периода и впоследствии разделилась на две основные формы, известные сегодня как подклассы бескилевых и новонёбных птиц. Бескилевые представляют относительно небольшую группу, куда относят страусообразных и скрытохвостых. Следующей группой, разошедшейся с остальными новонёбными птицами, стали представители надотряда Galloanserae, в состав которого включают гусеобразных и курообразных. Период, когда началось данное расхождение, является предметом научных споров — большинство специалистов считают, что веерохвостые получили своё развитие в меловом периоде, а ветвь Galloanserae отделилась от остальных новонёбных до начала мел-палеогенового вымирания. Тем не менее, у этой версии имеются и противники, полагающие, что расхождение могло произойти уже после (и благодаря) гибели большей части животного мира. Разногласие отчасти вызвано различными методами исследований:молекулярная генетика указывает на основную диверсификацию в середине мелового периода, а анализ окаменелостей свидетельствует о начале палеогена.
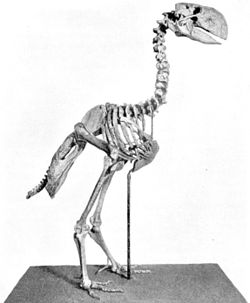
Скелет гасторниса
Древнейшей известной науке веерохвостой (настоящей) птицей является Archaeornithura meemannae, жившая около 130,7 млн лет назад на территории современного Китая.
Примерно 60 — 40 млн лет назад произошла интенсивная адаптивная радиациявеерохвостых птиц и возникновение большинство современных отрядов. Окаменелости птиц этого времени малочисленные и не дают представления о темпах и характере эволюции птиц в этот период. Птицы, известные из кайнозоя, вероятно, имеют общего предка выше уровня зубастых Hesperornithiformes и Ichthyornithiformes. Ископаемые новонёбные птицы современных отрядов известны преимущественно по фрагментарным отдельным остаткам из кампанских и маастрихтских отложений Монголии и Северной Америки. Они являются представителями таких отрядов, как гусеобразные, пеликанообразные, курообразные, трубконосые,ржанкообразные, гагарообразные. Также описан попугай из позднемеловых отложений США, хотя его групповая принадлежность остаётся спорной.
Бескилевые, вероятно, произошли от общего предка, отделившегося от ранних килевых форм, сохранивших палеогнатическое небо. Форма килевой кости современных страусов предполагает, что их дальние предки летали, и что не летающий образ жизни современных бескилевых птиц является вторичным явлением.
В большинстве отрядов птиц современные роды известны, начиная со среднего или позднего миоцена. Также многие из ныне живущих семейств птиц можно проследить по ископаемым останкам до позднего эоцена или олигоцена. Более ранние находки преимущественно настолько отличаются от современных родов, что установить их принадлежность к какому-либо семейству часто затруднительно.
Наряду с современными отрядами в кайнозое возник ряд семейств и отрядов птиц, по тем или иным причинам, впоследствии вымерших. Это гигантские не летающие бескилевые эпиорнисообразные и моаобразные, а также летающие бескиливые литорнитообразные. С палеоцена до плиоцена на территории Южной Америки обитали фороракосовые - вымершее семейство очень крупных хищных нелетающих птиц из отряда журавлеобразных. Ещё одну группу вымерших крупных нелетающих птиц составляют Gastornithidae из палеоцена и эоцена Северной Америки, Европы и Китая. Её представители ранее выделялись в самостоятельный отряд диатримообразные, но в настоящее время рассматриваются в качестве семейства в составе отряда гусеобразных.
Систематика
Первую попытку систематизации животных в IV веке до н. э. предпринял греческий учёный Аристотель — в своих сочинениях «О частях животных» и «О возникновении животных» он выделил всех известных ему птиц в «высший» род Ornithes. Несмотря на очевидное несовершенство этой системы, вплоть до второй половины XVII века новых попыток классификации животного мира не предпринималось. В 1676 году вышла в свет рукопись «Ornithologiae libri tres», написанная английским биологомФренсисом Виллоуби и уже после его смерти оформленная и опубликованная. Это первая известная работа, в которой авторы попытались классифицировать птиц на основании внешних морфологических признаков. В 1713 году была опубликована одна из первых классификаций птиц, сделанная английским биологом Джоном Рэем. Дальнейшие основы классификации птиц были заложены в 1758 году Карлом Линнеем в его «Системе природы», в которой он ввёл иерархические категории ибиноминальную номенклатуру при обозначении видов, используемые вплоть до настоящего времени. В системе Линнея все животные были разбиты на шесть основных категорий — классов, — одну из которых заняли птицы, или Aves (позднее появились категории и более высокого порядка). В настоящее время название Avesобычно употребляется по отношению к группе, включающей в свой состав всех потомков общего предка археоптерикса и современных птиц. Альтернативная филогенетическая систематика рассматривает группу Aves в качестве клады тероподных динозавров. Согласно теории происхождения птиц от динозавров, Aves и её сестринская клада Crocodilia (крокодилы) считаются единственными современными ветвями клады рептилий Archosauria.
К концу XIX века исследования Томаса Гексли, Михаила Мензбира, Макса Фюрбрингера, Ганса Гадова и др. также заложили основу для современных представлений о системе класса птиц и установили их филогенетические связи. В отличие от своих предшественников, для анализа родства видов они использовали большой по объёму сравнительно-анатомический материал.
В дальнейшем многие авторы по сути совершенствовали имеющуюся систему птиц.
Современная систематика
На протяжении длительного периода птицы рассматривались в качестве чётко очерченной группы, морфологически отделённой от других групп позвоночных. Однако после открытия и включения в состав птиц археоптерикса морфологические границы данной группы расширились. Тем не менее, ряд последователей системы Филокод, таких как Жак Готье, Луис Киаппи и др., под категорией Aves понимают лишь современных птиц, не включая туда группы, известные лишь по ископаемым останкам — археоптерикса, энанциорниса, конфуциусорниса и некоторых других. Эти учёные ввели новую кладу Avialae, или птиц в широком смысле, которая объединяет живущие в наше время виды и их ископаемых предшественников.
Все ныне живущие современные эволюционные группы птиц представляют собой монофилетическую группу Neornithes (веерохвостые птицы), которая противопоставляется ящерохвостым птицам — Saururae, или Archaeornithes. Веерохвостых птиц, в свою очередь, разделяет на две группы — Бескилевые иНовонёбные, которые обычно рассматриваются в ранге надотрядов, хотя в работах Лайвези и Зуси они рассматриваются как когорты.
Современные филогения и классификации
Современная филогения и классификация птиц до сих пор находится в стадии формирования. Сравнительный анализ анатомии птиц, ископаемых окаменелостей иДНК не привёл к единому мнению исследователей по этому вопросу.
К середине XX века существовал целый ряд сходных классификаций птиц, базирующихся преимущественно на данных сравнительной морфологии. Среди них наиболее распространённой была классификация Александра Ветмора (Wetmore, 1960), включавшая 27 современных отрядов. В ней автор разделял современных птиц на два крупных надотряда: Impennes (включавший пингвинов) и Neognathae (новонёбные, остальные современные птицы).
Традиционной из современных морфологических систем также считается классификация по Клементсу (Klements, 2007), претерпевшая три пересмотра. Согласно ей,веерохвостые птицы разделяются на 2 подкласса, объединяющие 33 отряда (6 из них только с ископаемыми формами) и 213 семейств (из них 42 — ископаемые). Существует классификация по Howard & Moore (2003), отличающаяся от системы Клементса выделением самостоятельных отрядов трёхпёрсткообразные(Turniciformes), удодообразные (Upupiformes) и Bucerotiformes.
В последние два десятилетия систематика и филогения птиц были значительно пересмотрены с широким внедрением различных методов молекулярного анализа. Первой классификацией птиц, основывающейся на использовании молекулярных биохимических данных, была классификация Сибли — Алквиста, созданная американскими учеными Чарльзом Сибли и Джоном Алквистом в 1970-х годах. В 1990 году авторами была опубликована работа «Филогения и классификация птиц» (Phylogeny and Classification of Birds), посвящённая филогении и эволюции птиц на основе ДНК-ДНК-гибридизации. Она стала фактической основой предложенной классификации. Однако классификация Сибли — Алквиста не стала общепринятой, морально устарев уже на момент своей публикации.
На сегодняшний день имеется целый ряд различных схем филогении современных птиц, полученных по результатам различных молекулярных и генетических исследований. Многие выводы на основании их результатов являются логичными, но ряд новых идей противоречит морфологическим и биогеографическим данным. Также филогении и классификации на основе результатов этих исследований, в зависимости от выбранных методик и маркеров, часто противоречат друг другу (напр.,Hackett et al., 2008; Pacheco et al., 2011). Преимущественно это связано с несовершенством применяемых методик и трактовок их результатов, конкуренцией научных школ, ошибками в выборе маркеров и т. п.. Последний обширный молекулярный анализ филогении птиц Prum et al. (2015) по результатам секвестирования ДНК охватывает собой 198 видов современных птиц, представляющих все основные линии, а также 2 вида крокодилов в качестве аутгруппы. В нем применялись Байесовский анализ и метод максимального правдоподобия, которые дали хорошо обоснованные и идентичные филогенетические деревья для всех основных линий птиц. Результаты анализа времени дивергенции совпадают с палеонтологической летописью, поддерживая крупную радиацию птиц вскоре после мел-палеогенового вымирания.
Наряду с этими двумя направлениями, развиваются компромиссные классификации, одновременно учитывающие последние достижения существующих молекулярных и морфологических классификаций птиц. Например, в 2001 году русским орнитологом Евгением Кобликом была предложена компромиссная система птиц, объединяющая традиционные морфологические представления и данные ДНК-ДНК-гибридизации. От классификации Ветмора она отличается широкой трактовкой страусообразных, выделением в отдельные отряды трёхперсток, фламинго, грифов Нового Света и рябков.
Международный союз орнитологов также использует собственную компромиссную классификацию по Gill & Wright (2006). Она представляет собой версию классификации по Howard & Moore, модифицированную данными ДНК-исследований птиц, обобщённых в работе «Avian Higher-Level Phylogenetics» (2003). Она содержит 40 отрядов, 239 семейств и 2 incertae sedis, 2282 рода птиц.
Кладограмма филогении современных птиц по Jarvis, E.D. et al. (2014) и Yury, T. et al. (2013)
| Традиционная классификация по Klements (2007) | Молекулярная классификация Sibley & Monroe (1990) |
| Подкласс Paleognathae — Бескилевые Отряд Казуарообразные (Casuariiformes) Отряд Кивиобразные (Apterygiformes) Отряд Нандуобразные (Rheiformes) Отряд Страусообразные (Struthioniformes) Отряд Тинамуобразные, или скрытохвостые (Tinamiformes) † Отряд Эпиорнисообразные (Aepyornithiformes) Подкласс Neognathae — Новонёбные Отряд Аистообразные, или голенастые (Ciconiiformes, Gressores) Отряд Буревестникообразные, или трубконосые (Procellariiformes, Tubinares) Отряд Воробьинообразные (Passeriformes) Отряд Гагарообразные (Gaviiformes) † Отряд Гесперорнисообразные (Hesperornithiformes) Отряд Голубеобразные (Columbiformes) Отряд Гусеобразные (Anseriformes) † Отряд Диатримообразные (Diatrymiformes или Gastornithiformes) Отряд Дятлообразные (Piciformes) Отряд Журавлеобразные (Gruiformes) † Отряд Ихтиорнисообразные (Ichthyornithiformes) Отряд Козодоеобразные (Caprimulgiformes) Отряд Колибриобразные (Trochiliformes) Отряд Кукушкообразные (Cuculiformes) Отряд Курообразные (Galliformes) Отряд Пеликанообразные, или веслоногие (Pelecaniformes, Steganopodiformes) Отряд Пингвинообразные (Sphenisciformes) Отряд Поганкообразные (Podicipediformes) Отряд Попугаеобразные (Psittaciformes) Отряд Птицы-мыши (Coliiformes) Отряд Ракшеобразные (Coraciiformes) Отряд Ржанкообразные (Charadriiformes) Отряд Рябкообразные (Pteroclidiformes) Отряд Совообразные (Strigiformes) Отряд Соколообразные, или дневные хищные птицы (Falconiformes) Отряд Стрижеобразные, или длиннокрылые (Apodiformes) Отряд Трогонообразные (Trogoniformes) Отряд Туракообразные (Musophagiformes) Отряд Фламингообразные (Phoenicopteriformes) | Отряд Гусеобразные Отряд Стрижеобразные Отряд Bucerotiformes — Птицы-носороги включены в ракшеобразных Отряд Ciconiiformes Аистообразные Ржанкообразные Соколообразные Гагарообразные Пеликанообразные Фламингообразные Поганкообразные Буревестникообразные Пингвинообразные Отряд Coliiformes Отряд Columbiformes Отряд Coraciiformes Отряд Cuculiformes Отряд Galbuliformes — Якамаровые включены в дятлообразных Отряд Курообразные Отряд Журавлеобразные Отряд Musophagiformes — Турако включены в кукушкообразных Отряд Воробьинообразные Отряд Дятлообразные Отряд Попугаеобразные Отряд Strigiformes Совообразные Козодоеобразные Отряд Struthioniformes Страусообразные Нандуобразные Казуарообразные Кивиобразные Отряд Тинамуобразные Отряд Trochiliformes — Колибри включены в стрижеобразных Отряд Трогоновые Отряд Turniciformes — Трёхперстко́вые включены в журавлеобразных Отряд Удодообразные |